11
Die Totenkopfaffenkolonie der Universität Kassel
Ich habe in diesem Buch bisher
nur über unsere Ergebnisse an Affen berichtet, die paarweise oder in kleinen
Familienverbänden gehalten werden (Kapitel 6 bis Kapitel 10). Hier und in den
folgenden Kapiteln stelle ich Ergebnisse von Arten vor, die geselliger leben, bei denen
Männchen und Weibchen toleranter zu Individuen des eigenen Geschlechtes sind. Man
kann und sollte sie daher in größeren Sozialgruppen halten. Leider gilt auch für die
Individuen dieser Arten, dass - wie ich es in Kapitel 1 betont habe - der Artgenosse der
Hauptkonkurrent des jeweiligen Individuums ist. Insofern treten auch in den
Sozialverbänden dieser geselliger lebenden Affen massive Konflikte auf. Da Affen für
das Austragen ihrer Konflikte nur ihre Zähne zur Verfügung haben, kommt es
zwangsläufig auch zu Verletzungen, geringerer oder massiverer Qualität. Bei den
Auseinandersetzungen handelt es sich eigentlich immer um Dominanzprobleme.
Man muss sich diese Gruppen als Summe von Zweierbeziehungen vorstellen,
wobei natürlich jedes Gruppenmitglied gleichzeitig mehrere Zweierbeziehungen
eingehen und ertragen muss. Jedes Individuum in einer Zweierbeziehung möchte
das dominantere sein. Unter Freilandbedingungen entscheiden die Verlierer
solcher Dominanzkämpfe, ob sie dennoch in der Gruppe bleiben und sich mit
einer weniger dominanten Position innerhalb des Sozialverbandes zufrieden
geben oder ob sie die Gruppe verlassen. Da das Verlassen der Gruppe unter
natürlichen Bedingungen ein erhebliches Lebensrisiko bedeutet, die Selektion ist
hier unbarmherzig, hat sich in der Evolutionsgeschichte das Ausharren in
der Gruppe als die überlegene Strategie herausgestellt. Nur in dem größeren
Sozialverband kann das Individuum seine „Darwinian fitness“ (vgl. Kapitel 2)
mehren.
Auseinandersetzungen und besonders auch schwere Verletzungen kosten Energie, die
unter natürlichen Bedingungen nur begrenzt zur Verfügung steht, insofern müssen sich
in der Evolutionsgeschichte Mechanismen entwickelt haben, die Auseinandersetzungen
minimieren. Diese Regeln des friedlichen Zusammenlebens lassen sich verkürzen auf die
Faustregel: „Der mir Vertraute ist der Gute, der mir Fremde ist der Böse.“ Leider gilt
dies nur für das friedliche Zusammenleben der Gruppen. Der vertraute Partner ist
nämlich als Sexualpartner unattraktiv. Unter natürlichen Bedingungen wechseln daher
regelmäßig die Männchen, sie verlassen die Gruppe und suchen andere Sozialverbände
auf. Freilich ist dies bei freilebenden Gruppen mit großen Verlusten verbunden. In
Menschenobhut ist hier der verantwortliche Halter gefragt, entsprechende Prozesse zu
simulieren. Die verschiedenen Arten, über die ich berichte, zeigen uns verschiedene
Wege der Realisation und Lösung dieser Probleme. Diese zu belegen, war das
Forschungsanliegen, das mich in meiner jahrzehntelangen Arbeit ausgelastet und
belastet hat.
Unter den Luxusbedingungen in Menschenobhut, ohne die Gefahren des Verhungerns
und ohne den Druck durch Predatoren, können die Individuen freilich nicht
entscheiden, ob sie die Gruppe lieber verlassen würden, diese Entscheidung
übernimmt der verantwortliche Halter. Ich hatte in den bisherigen Kapiteln immer
wieder herausgestellt, dass ich das Todesrisiko vermeiden wollte und daher
aus dem Familienverband verdrängte Individuen stets separiert habe. Diese
Halterstrategie ist bei in größeren Gruppen lebenden Primaten nicht anwendbar, da
dann in letzter Konsequenz nur Einzelhaltung möglich wäre, was zweifellos
nicht artgemäß ist (und auch keine Untersuchungen zum Sozialverhalten
ermöglichte). Insofern habe ich alle bedrängten Individuen dann im Sozialverband
gelassen, wenn der Konflikt entschieden schien und der Unterlegene sich mit
seiner unterlegenen Stellung abgefunden hatte. Nur die Individuen wurden
abgetrennt, die nach verlorenem Kampf auch weiterhin Ziel von massiven
Aggressionen waren, sich also unter natürlichen Bedingungen zweifellos für das
Verlassen der Gruppe entschieden hätten. Die übrigen durften im Sozialverband
bleiben.
Freilich sahen die Verlierer der Konflikte häufig nicht attraktiv aus, sie hatten im
besten Fall nur Haare verloren, im schlimmsten Fall erhebliche Verletzungen, die
ausheilen mussten. Den menschlichen Beobachter rühren diese Individuen an. Wir
denken, man müsse sie behandeln, sie separieren und gesundpflegen. Eine
dementsprechende Entscheidung ist aber falsch. Eine Entfernung eines Individuum aus
dem Sozialverband, um es später wieder zu integrieren, gelingt nur, wenn das
separierte und verletzte Individuum der Sieger des Konfliktes war, der Sieger
kehrt mehr oder weniger problemlos in den Sozialverband zurück. Die übrigen
Gruppenmitglieder erinnern sich an ihn und ordnen sich unter. Separiert man hingegen
den Verlierer, dann stürzen sich nach der Reintroduktion alle Gruppenmitglieder auf
ihn, um bereits bestehende Dominanzverhältnisse zu festigen, mit der Konsequenz,
dass der Verlierer nun eine noch schlechtere Position hat (bzw. noch schwerer verletzt
ist) als vor der Separation.
Die richtige und bewährte Entscheidung, verletzte Individuen im Sozialverband zu
belassen, lässt sich freilich nur durchhalten, wenn sich nicht ständig menschliche
Primaten über den Halter beschweren. Sie ist also kein guter Rat für kommerziell zur
Schau gestellte Primaten.
Als erstes möchte ich über unsere Haltungserfahrungen mit
Totenkopfaffen
berichten. Unsere ersten zehn Totenkopfaffen, drei Männchen und sieben
Weibchen, importierten wir zum Versuch des Aufbaus einer Zuchtkolonie am
27.09.1975 aus British Guayana, Abflughafen Georgetown. Wir ernährten sie
erfolgreich mit unserer Diät (am Morgen einen vitaminisierten eiweißhaltigen
Brei, mittags Kartoffeln, Zwieback und Eier, nachmittags Obst und Gemüse
nach Angebot) und hatten offensichtlich auch Glück, alle Tiere überlebten die
Strapazen des Transports und durchliefen erfolgreich unsere Quarantäne. Bevor ich
detaillierte eigene Ergebnisse vorstelle, möchte ich den damaligen Wissenstand
wiedergeben.
Für Totenkopfaffen werden Gruppen bis 300 - 500 Tiere
berichtet,
doch ist es äußerst schwierig, die einzelnen Mitglieder der Gruppe
voneinander zu unterscheiden, bzw. überhaupt Gruppen kontinuierlich zu
beobachten.
Zum Zeitpunkt des Beginns unserer eigenen Untersuchungen lagen nur zwei
Feldstudien vor, beide an kleinen Sozialverbänden.
R. W. Thorington ([224]) berichtet von seiner Feldforschung in Kolumbien, dass alle
Tiere nachts gemeinsam schlafen. In der Morgendämmerung wachen sie auf und
bewegen sich dann in kleinen Untergruppen zu den Futterbäumen. Er konnte
drei typische Untergruppen voneinander unterscheiden; getrennt wandern
nämlich erwachsene Männchen, erwachsene Weibchen und Junge tragende
Weibchen; ältere Jungtiere schließen sich meist den Weibchenuntergruppen
an. Die einzelnen Untergruppen halten sich nie lange in einem Baum auf,
sondern wechseln häufig von Baum zu Baum; auf dem Weg suchen sie dann
nach Insekten und Spinnen. Niemals „plündern“ sie den Baum völlig.
Thorington beobachtete, dass eine Untergruppe nach der anderen denselben
Baum besuchte, beziehungsweise dass die Tiere mehrfach zum selben Baum
zurückkehrten.
Diese deutliche Untergruppenaufspaltung konnten die Baldwins bei ihren
Totenkopfaffen in Panama ([7]) nicht feststellen, wenngleich sich auch hier die
erwachsenen Männchen am Rand der Gruppe aufhielten. In der Regel leben
Totenkopfaffen in der mittleren Waldschicht, gelegentlich werden sie aber auch am
Boden oder im Unterholz angetroffen. In allen drei genauer untersuchten Gruppen war
jeweils die Zahl der Kinder und Jungtiere am größten, die Zahl der erwachsenen
Weibchen übertraf die Zahl der erwachsenen Männchen. Totenkopfaffen legen 2,5 bis
4,2 km am Tag zurück. Streitigkeiten in der Gruppe sind selten zu beobachten;
entdecken die Tiere bei ihrem Herumstreifen eine neue Futterquelle, stürzen sie oft alle
gleichzeitig herbei und versuchen, die Früchte zu erlangen; dabei klettern sie
auch übereinander. Die Wohngebiete verschiedener Gruppen überlappen, feste
Eigenbezirke (Territorien) gibt es also nicht. Auffällig war, dass die Juvenilen nicht
spielten.
Weitaus genauere Beobachtungen lagen von einer unter halbnatürlichen Bedingungen in
einem Waldgebiet von 1,6 Hektar im südlichen Florida, dem Monkey Jungle, in der Nähe
von Miami
gehaltenen Kolonie vor. Hier untersuchte wiederum Baldwin gründlich deren Sozialleben
([4], [5], [6]). Er belegte, dass die Gruppen in Untergruppen aufgespalten sind, außerdem
sammelte er umfangreiche Kenntnisse über das Fortpflanzungsverhalten und die Entwicklung
der Jungtiere. Hierdurch schuf er Grundlagen für unser Wissen über freilebende
Totenkopfaffen.
Gerade im Hinblick auf die „Rangordnungsdebatte“, auf die ich im Kapitel 14
ausführlich eingehen werde, möchte ich noch einen Befund von Baldwin erwähnen:
„There were no leaders or control animals in the troop, and social hierarchies were
absent except among the troop’s 4 adult males during the mating season.“ ([6], Seite
23).
Auch in Deutschland hat eine Arbeitsgruppe um Detlev
Ploog
in München am dortigen Max-Planck-Institut für Psychiatrie eingehend das Verhalten
der Totenkopfaffen erforscht. Nahezu die gesamte Pionierarbeit zum Verhalten der
Totenkopfaffen wurde hier geleistet.
Detlev Ploog war auch der erste, der darauf hinwies, dass das Verhaltensprofil jedes
Gruppenmitgliedes unterschiedlich ist. ([159], [154]). Diese Aussage kommt uns heute,
50 Jahre später, sicherlich selbstverständlich vor, doch war sie damals, als noch viele
Forscher glaubten, dass Affe gleich Affe und erst recht Totenkopfaffe gleich
Totenkopfaffe sei, eine aufsehenerregende Erkenntnis und Ausgangspunkt für
nahezu alle späteren Untersuchungen an Totenkopfaffen. Detlev Ploog war vor
allem an der Kommunikation der Totenkopfaffen (und der Primaten generell)
interessiert. Ihm verdanken wir hier grundlegende Arbeiten (u. a. [156], [157],
[158]).
An seiner Kolonie erarbeitete auch Sigrid Hopf in mühevoller Arbeit den
ersten vollständigen Verhaltenskatalog für Totenkopfaffen ([90], [91], [92],
[95]), der auch Grundlage für unsere eigenen Arbeiten war. Beiden verdanken
wir auch Erkenntnisse zum angeborenen Verhalten von Totenkopfaffen (u. a.
[97]).
Sigrid Hopf führte gemeinsam mit Michael Herzog hierzu weitere Experimente mit
isoliert aufgezogenen Totenkopfaffen durch ([96]).
Detlev Ploog beschrieb und deutete auch als erster das „genitale
Imponieren“ (Zurschaustellung der Geschlechtsteile). ([155], [154] Dabei
nähern sich die Tiere einander mit erigiertem Penis bzw. erigierter
Klitoris und abgewinkeltem Bein. Ploog erkannte, dass dies ein deutliches
Zeichen für Dominanzbeziehungen („Rangordnung“) unter Totenkopfaffen
ist.
Niemals beobachtete er das wechselseitige Imponieren zwischen zwei
Männchen.
„Diese Verhaltensweise erwies sich als ein sehr wirksames soziale Signal, das wesentlich
zur Konstituierung einer hierarchischen Ordnung der Gruppe beitrug.“ Ploog fand
aber auch bereits in seinen ersten Arbeiten, dass „das herkömmliche Konzept einer
linearen Rangordnug ... sich für die Darstellung der Gruppe als unzureichend“ ([154],
Seite 107) erweist.
Das genitale Imponieren einem anderen Tier gegenüber mag bei adulten Männchen
ein eindeutiges Zeichen dafür sein, dass der imponierende Totenkopfaffe auch wirklich
überlegen ist. Für die Beschreibung des Sozialgefüges der Gruppe reicht aber diese eine
Verhaltensweise nicht aus. Einmal wechseln, wie es die Baldwins wiederum beobachten
konnten, Dominanzbeziehungen zwischen erwachsenen Männchen schnell und häufig,
zum anderen lassen sich bei Weibchen in der Regel keine klaren Dominanzbeziehungen
erkennen.
Wie bereits betont, belegen Beobachtungen im Freiland und in Menschenobhut
jedoch, dass Männchen vor allem mit Männchen und Weibchen vor allem mit
Weibchen engen Körperkontakt halten. Totenkopfaffengruppen zerfallen also in
gleichgeschlechtliche Untergruppen. Erwachsene Weibchen stehen im Mittelpunkt der
Gruppe, erwachsene Männchen hingegen leben am Rande und unterhalten auch
untereinander häufig nur geringen Kontakt. Nun darf man sich dies nicht so vorstellen,
dass die erwachsenen Weibchen eine geschlossene Untergruppe bilden, vielmehr
können wir in größeren Sozialgruppen stets mehrere Weibchenuntergruppen
unterscheiden.
Diese Annahme wird durch Versuche von Lynn Fairbanks ([54]) gestützt. Sie
untersuchte eine aus zwei Männchen und sechs Weibchen bestehende Gruppe und
fand, dass die Weibchen zwei geschlossene Untergruppen von jeweils drei Tieren
bildeten, wobei in jeder dieser Untergruppen zwei Weibchen besonders enge
Beziehungen unterhielten. Durch ein interessantes Experiment konnte sie zeigen, dass
die Untergruppen tatsächlich geschlossen sind. Sie teilte den Käfig in zwei Bereiche
und trennte so die Untergruppen jeweils mit einem der Männchen räumlich
voneinander ab. In einem ersten Experiment setzte Lynn Fairbanks dann ein Weibchen
der Untergruppe B in den Käfig der Untergruppe A. Das Weibchen fand
keinen Anschluss an die Weibchengruppe, sondern konnte nur Beziehungen zu
dem Männchen der Untergruppe A aufnehmen, bildete mit diesem also eine
gemischtgeschlechtliche Untergruppe. Nach der im zweiten Versuchsabschnitt
durchgeführten Wiedervereinigung der acht Totenkopfaffen fand dieses Weibchen
wieder Anschluss an seine alte Weibchenuntergruppe. der Kontrollversuch
(ein Weibchen der Untergruppe A zur Untergruppe B) erbrachte das gleiche
Ergebnis.
Totenkopfaffen gehen also offensichtlich keine gemischtgeschlechtlichen Paarbindungen
ein, wie wir sie in den Kapiteln 9 und 10 für die Spring- und Nachtaffen berichtet
haben. Durch die vergleichenden Beobachtungen von William A. Mason, über die ich
ausführlich in Kapitel 9 berichtet habe, konnten die Partnerpräferenzen bei
Totenkopfaffen eindeutig durch umfangreiche Experimente belegt werden. Er
hielt seine Totenkopfaffen stets paarweise zusammen, so dass beide Tiere
völlig miteinander vertraut waren. Setzte er nun zu einem solchen Paar ein
fremdes Weibchen hinzu, dann nahmen beide Weibchen engen Kontakt auf, der
Fremdling wird also sogar dem vertrauten Männchen vorgezogen. Das enge
Verhältnis der Weibchen untereinander ist also die wichtigste Komponente des
Sozialgefüges.
Die soziale Organisation der Gruppe wird aber auch durch den jährlichen
Fortpflanzungszyklus beeinflusst. In der natürlichen Umgebung nämlich bekommen
alle Weibchen im gleichen Zeitraum ihre Kinder. Entsprechend können wir das Jahr
unterteilen in eine Paarungszeit, eine Geburtensaison und eine Aufzuchtszeit. Während
der Paarungszeit werden die sonst nur am Rand der Gruppe lebenden Männchen sehr
aktiv und erregbar und kämpfen auch miteinander. Darüber hinaus nehmen sie zudem,
zumindest in einigen Lebensräumen und in einigen Kolonien in Menschenobhut,
erheblich an Gewicht zu und suchen die Nähe der Weibchen, werden aber auch häufig
von mehreren erwachsenen Weibchen gemeinsam abgewehrt. Ein Werbeverhalten - wie
wir es für den Gehaubten Kapuziner beschreiben werden (vgl. Kapitel 13) - wurde
bisher nicht beobachtet.
Totenkopfaffen werden in der Regel nachts geboren. In den ersten Lebenswochen
zeigen die Mütter und andere Gruppenmitglieder wenig Interesse an dem Kleinkind,
das bereits vom ersten Lebenstag an auf dem Rücken der Mutter getragen
wird. Dieser Lebensabschnitt scheint jedoch wichtig für die Entstehung der
späteren engen Mutter-Kind-Bindung zu sein. Matt J. Kessler und Hector S.
Martinez vom „Karibischen Primatenzentrum“ auf Puerto Rico konnten nämlich
insgesamt dreimal beobachten, dass ein Weibchen zusätzlich zu dem eigenen
Jungen ein zweites Kind erfolgreich großzog, das am selben Tag wie das eigene
geboren war. Jeweils erfolgte der Wechsel zu Ende der ersten Lebenswoche.
Versuche, den Müttern ihre Jungen zurückzugeben, scheiterten. Die drei
„guten“ Mütter belegten zudem, dass Totenkopfaffen ohne weiteres in der
Lage sind, auch Zwillinge großzuziehen, obwohl Zwillingsgeburten bisher für
Totenkopfaffen noch nicht berichtet worden sind. Die Gewichtsentwicklung
verlief jedenfalls normal, wenngleich etwas langsamer als bei Einlingen.
([111])
In der dritten und vierten Woche wird das Junge beweglicher und ruft die
Aufmerksamkeit anderer Weibchen hervor, die sich dann auch als „Tanten“ an dem
Transport des Jungen beteiligen können. In der fünften bis zehnten Lebenswoche
schließlich beginnt das Junge, gelegentlich den Rücken der Mutter zu verlassen, die
Umgebung zu erkunden und erste feste Nahrung zu sich zu nehmen. Ab der achten
Woche bis zum vierten Monat werden Mutter-Kind-Kontakte immer seltener.
Während des fünften bis zum zehnten Monat wird das Junge zunehmend unabhängig
von der Mutter.
Soziales Spiel beginnt nach den Untersuchungen im Labor in München und unter
halbnatürlichen Bedingungen in Florida zu Ende des zweiten Lebensmonats. Nun
balgen die kleinen Totenkopfaffen oft miteinander. Im ersten Jahr beteiligen sich
sowohl männliche als auch weibliche Kinder an den Kampfspielen. Im zweiten
Lebensjahr suchen die jungen Weibchen jedoch immer häufiger den Kontakt zu den
erwachsenen Weibchen der Gruppe und stellen bei Erreichen der Geschlechtsreife das
Spielen ein, während Männchen auch noch als Jungerwachsene häufig bei
Kampfspielen zu beobachten sind.
Die Kindheit der Totenkopfaffen dauert etwa bis zum Ende des elften Lebensmonats,
die Jungtierphase vom 12. bis 30. Lebensmonat. Nun sind die Weibchen erwachsen, die
Männchen dagegen durchlaufen noch eine Reifungsphase bis zum Ende des vierten
oder zur Mitte des sechsten Lebensjahres.
Auffällig bei den Totenkopfaffen ist eine besondere und typische Ruhestellung. In der
Regel legen sie nämlich wie unsere einheimischen Bilche den Kopf nach unten und den
Schwanz von vorne über die Schulter.
Nur bei hohen Temperaturen oder nach Zeiten hoher Bewegungsaktivität liegen sie auf
dem Bauch mit nach den Seiten fortgestreckten Armen und Beinen, wohl um sich
abzukühlen. Möglicherweise im Zusammenhang mit der Temperaturregulation steht
auch ein zweites auffälliges Verhalten: Totenkopfaffen „waschen“ sich mit Harn. Sie
harnen auf die Hand und verreiben den Harn mit der Fußfläche, wodurch Hand- und
Fußsohle angefeuchtet werden. Manchmal reiben sie auch anschließend den
Schwanz ein. Häufiger können wir noch das Einreiben des Schwanzes mit Futter
beobachten; vielleicht erhalten so Totenkopfaffen, die unter Freilandbedingungen
im selben Gebiet Nahrung suchen, einen „Gruppengeruch“. Zum Markieren
dient das Waschen mit Harn wahrscheinlich nicht. Entsprechendes Verhalten
habe ich für den Nachtaffen (Kapitel 10) angegeben und werde es auch für
den Kapuzineraffen berichten (Kapitel 13), bei den nachtaktiven Halbaffen
Galago garnettii wurde es von mir experimentell untersucht (vgl. Kapitel
3).
Aber nun zurück zu unseren eigenen Totenkopfaffen. Von den zehn 1975 importierten
Tieren starben zwei Weibchen 1976. Zwei der drei Männchen mussten wir aus dem
Sozialverband entfernen, wir hielten sie in Sicht-, aber ohne Gitterkontakt. 1977
züchtete die Gruppe erstmals. Alle fünf Weibchen wurden mehr oder weniger
gleichzeitig schwanger, vier Weibchen (Mara, Toxa, Anga und Eda) zogen ihre Jungen
auch auf, das fünfte Weibchen (Dagmar) starb, ebenso wie sein Junges, nach einem
Kaiserschnitt. Diese Jungtiere wuchsen ohne helfende „Tanten“ heran. Das
Tantenverhalten ist somit nicht immer - wie das Tragen der jungen Spring- und
Nachtaffen durch den Vater - zu beobachten. Die Zucht- und Aufzuchtserfolge
wurden in dieser Gruppe zur Regel. Eda zog bis zu ihrem Tod (08.10.1981)
fünf Junge groß, Toxa (gest. 28.12.1984) sieben Junge, Anga (bis zu ihrer
Abgabe an andere Halter 1984) drei Jungtiere und Mara bis zu ihrem Tod
am 21.07.1990 sogar acht Junge (zudem drei Fehlgeburten). Mara war bei
der Geburt ihres letzten Kindes, einem Männchen, Janus (Vater: Tim (s.
u.)), am 24.10.1989, mindestens 16 Jahre alt. Eines der Gründermännchen,
Bruno, ließ ich am 09.06.1996 hochbetagt einschläfern, 21 Jahre nach Import.
Totenkopfaffen können also alt werden. Bereits 1980 wurde bei uns das erste Kind
der zweiten und 1985 das erste Kind der dritten Generation geboren und
aufgezogen.
Gemeinsam mit meiner Mitarbeiterin Annette Klaiber-Schuh ([270]) wertete ich
1995 auch bei den Totenkopfaffen die Reproduktionsdaten unserer Kolonie
aus. Insgesamt wurden bis zum Zeitpunkt der Auswertung 103 Kinder (nur
Einlinge) geboren. Die Überlebensrate war hoch. 74 % der Neugeborenen
überlebten die erste Lebenswoche, von den Überlebenden erreichten dann
88 % zumindest das erste Lebensjahr. Erstgebärende Weibchen (n = 17)
waren zwar weniger erfolgreich als mehrfachgebärende, doch überlebten 59
% der Kinder die erste Lebenswoche. Die zweitgeborenen Jungtiere hatten
jedoch die gleichen Chancen wie die Kinder der übrigen mehrfach gebärenden
Weibchen, 89 % erreichten das Ende der ersten Lebenswoche. Im Gegensatz zu
den bisher besprochenen Affenarten können wir bei den gesellig lebenden
Totenkopfaffen auch Informationen zur Geschlechtsreife geben. Das mittlere Alter der
erstgebärenden Weibchen war 3 Jahre 9 Monate (± 4 Monate). Fünf der
Weibchen waren zum Zeitpunkt der Geburt des ersten Kindes exakt drei Jahre
alt.
Der Geburtenabstand hängt nicht von dem Aufzuchtserfolg ab, er betrug
417 ± 22 Tage bei Kinder tragenden Weibchen (n = 43) und 417 ± 22 Tage
bei Weibchen, die das Kind verloren hatten (n = 21). Eine Saisonalität war
bei den Geburten in der Kolonie auffällig, die meisten Kinder wurden im
September und Oktober geboren, unabhängig von der Haltungsdauer in
unserer Kolonie und unabhängig von der Vorgeschichte (Wildfang oder
Nachzucht).
Das Geschlechterverhältnis Männchen zu Weibchen war 1 : 0,97 bei allen Jungtieren,
1 : 1 bei den überlebenden Kindern.
Der Hauptträger der Kinder war die eigene Mutter. Soweit Fremdelternpflege
überhaupt beobachtet werden konnte, war das Tragtier in der Regel entweder eine
Schwester des Kindes oder der Mutter. Vertrautheit (beruhend auf Verwandtschaft)
reduzierte die sexuelle Attraktivität. Der männliche Sexualpartner wurde außerhalb
des vertrauten sozialen Umfeldes gewählt. In der Gruppe geborene Kinder wurden
Mitglieder der Untergruppe der Mutter. Weibliche Kinder blieben in dieser
Untergruppe, männliche Kinder dagegen verließen die Untergruppe und fanden soziale
Kontakte hauptsächlich zu anderen Männchen.
1978/1979 ermöglichte ich es Jens Steffen, im Rahmen seiner
Staatsexamensarbeit
unsere Totenkopfaffen (vgl. Tabelle 11) zu beobachten.
Tabelle 11.1: Familiengruppe Saimiri sciureus 1978/1979
Vor Beginn seiner Untersuchung waren Bimbo und Bino separiert, sie hatten Sicht-
aber keinen Gitterkontakt, später wurden sie mit Gitterkontakt zur Familiengruppe
gehalten.
Bei seinem Verhaltenskatalog folgte er Hopf ([95]).
Zu Beginn seiner Untersuchung hatte Toxa keinen Kontakt zu den anderen
adulten Weibchen, drängte dann aber Mara aus der Weibchenuntergruppe,
so dass diese nur noch Kontakt zu den adulten (Bruno/Bino) und
juvenilen (Maro/Arno) hielt bzw. halten konnte. Steffen beobachtete kein
Tantenverhalten. „Interessant allerdings ist, dass Mara’s Junges einige Male von
den juvenilen Männchen Maro und Arno getragen wurde. ... Einmal konnte
ich
beobachten, dass Maro das Kind von Anga trug.
Ich
konnte darüber hinaus feststellen, dass die juvenilen Weibchen Mona und Erna
auffällig wenig Kontakt zu ihren Müttern hatten. Lediglich einmal war zu sehen, dass
Erna versuchte, Eda’s Junges zu greifen und wenig später auch für Sekunden zu
tragen.“ (Seiten 30 - 31). Steffen beobachtete bereits ab dem 13. Lebenstag das
Herunterklettern von der Mutter, doch suchten die Kinder in vermeintlichen
Gefahrensituationen die Mütter auch im Alter von neun Monaten auf. Bruno hatte
kaum Kontakte zum Rest der Gruppe, doch schien er diese zu verteidigen. „Immer
wenn eine fremde Person den Raum betrat, in welchem unsere Gruppe untergebracht
war, begann Bruno im Käfig hin- und herzuspringen, dabei am Gitter rüttelnd und
laute Schreie ausstoßend.“ (Seite 33) Steffen fasst die Ergebnisse seiner Untersuchung
zusammen:
„In der vorliegenden Arbeit wird aufgezeigt, dass wir in der Saimiri sciureus Gruppe
vier verschiedene Verhaltensprofile erkennen können. (1) Das Verhalten der adulten
Männchen, (2) das Verhalten der adulten Weibchen, (3) das Verhalten der juvenilen
Männchen und (4) das Verhalten der juvenilen Weibchen. ... Soziale Kontakte der
Männchen sind vornehmlich Spielkontakte, die der Weibchen beschränken sich meist
auf Kontaktsitzen.“(Seiten 35 - 36)
In der Folgezeit vergesellschafteten wir Bimbo mit Mona und Erna, Erna erfreute
uns am 05.10.1980 mit einem jungen Männchen (Bert) und am 02.09.1981
mit einem jungen Weibchen (Berit). Die restlichen Weibchen beließen wir in
der Gruppe, die jungen Männchen wurden für verschiedene Fragestellungen
entnommen oder als halbjährige Jungtiere an Privatleute (über den Tierhandel)
abgegeben.
Im November 1983 streckte (nach einer der Versionen, die ich gehört habe) im
Flughafen Frankfurt einer von vier Totenkopfaffen, die nach Deutschland geschmuggelt
wurden, seine Hand aus der Kiste, die Tiere wurden beschlagnahmt (die andere
Version: nach Zielfahndung). Wir erklärten uns bereit, sie zu übernehmen. In Kassel
stellten wir fest, es seien bestimmt andere Tiere als unsere bisherigen Totenkopfaffen.
Wir bestimmten sie als bolivianische Totenkopfaffen (Saimiri boliviensis), drei der
Tiere (Albo, Rika und Gesa) lebten dann jahrelang in unserer Primatenstation,
züchteten aber nicht.
Die richtige Zuordnung der Totenkopfaffen ist für alle mit Totenkopfaffen arbeitenden
Forscher ein großes Problem. Dabei wurden sie von den „Autoritäten“ regelrecht im
Stich gelassen, wie R. W. Cooper 1968 ausführte ([26]). Nach ihm wurden
Totenkopfaffen erstmals 1958 für biomedizinische Forschungsprojekte genutzt und zu
Tausenden
in die Laboratorien der westlichen Welt exportiert, sie sollten dort ein billigerer und
ungefährlicherer Ersatz für die Versuchsaffen des Genus Macaca sein. Die Forscher
erkannten sehr schnell, dass Totenkopfaffe nicht gleich Totenkopfaffe ist, doch „much
of the primate taxonomic literature is in a state of confusion, such that it is often
impossible to determine correct scientific nomenclature“([26], Seite 3). In seiner
Verzweiflung wandte er sich an Hershkovitz und erhielt 1966 den persönliche Rat „that
investigators using squirrel monkeys employ the binominal Saimiri sciureus.
Provenance, if known, and a good photograph of the entire animal with closeups of
front and side of head should be included in the paper. The author would do well to
give sex, age, weight and, if possible, some standard external measurements. If the
author believes two or more kinds of squirrel monkeys entered into his experiment, he
should describe the distinguishing characters.“ ([26], Seiten 11 -12). Cooper zumindest
befolgte den Rat und bildete die drei Hauptversuchstierarten (Totenkopfaffen
aus (1) Kolumbien, (2) Peru und (3) British Guayana) in seinem Beitrag
ab.
An der Zuordnungsdiskussion konnten/mussten wir uns dann beteiligen. Cooper
diskutiert auch die Bestimmung der Art nach der Form der Augenböden als
„romanisch“(rund) oder „gotisch“ (spitz). Er empfiehlt, zusätzlich jeweils den Ort
anzugeben, von dem die Tiere abgeschickt worden sind. (Dieser Aufforderung sind wir
gefolgt, s. o., s. u.)
In Vorbereitung auf den Umzug in die neue Primatenstation bildeten wir am
09.11.1984 zwei neue Sozialgruppen der Totenkopfaffen aus British Guayana, wir
verpaarten Bert mit sechs ihm nicht vertrauten Nachzuchtweibchen der Bruno-Gruppe
und introduzierten Berit in die Wildfang-Restgruppe. Zu diesem Zeitpunkt lebten
Bruno und Mara ununterbrochen neun Jahre gemeinsam zusammmen. Dennoch nahm
Berit sofort positiv soziale Kontakte zu Mara auf, beide Weibchen präferierten
einander vor Bruno.
Ein wichtiges Nebenergebnis dieser Neuverpaarungen war, dass nun (vorübergehend)
auch Geburten zur „falschen“ Zeit auftraten, so wurden 1985 Jungtiere im April und
Mai geboren, wofür der Umzug in die neue Primatenstation nicht verantwortlich
sein konnte, wurden die Jungtiere offensichtlich nach Verpaarung noch in der
alten Station gezeugt. Ich vermute daher, dass diese neue Konstellation das
„Geheimnis“ ist, warum gerade in großen Kolonien, in denen zwangsläufig
häufig neue Gruppen gebildet werden, Totenkopfaffen keine jahreszeitlichen
Übereinstimmungen im Fortpflanzungsgeschehen zeigen. Werden sie jedoch nach dem
Import stets zusammengelassen, wie in kleineren Kolonien üblich, bzw. haben sie
Zugang zu Außengehegen (und damit zu jahreszeitlichen Schwankungen im
Klimaverlauf) behalten sie den Jahreszyklus bei bzw. kehren zum Jahreszyklus
zurück.
In der neuen Primatenstation beobachtete 1985 Doris Merz, die
bereits in der alten Station Totenkopfaffen über vier Jahre beobachtet
hatte, im Rahmen
ihrer Diplomarbeit
unsere drei Totenkopfaffengruppen. „Erste Beobachtungen (an den drei Sozialgruppen)
führten zu dem überraschenden Ergebnis, dass diese Befunde (die Merz bereits in den
Jahren zuvor erhoben hatte) sich bei nur einer (Unter-)Art des Totenkopfaffen
verifizieren lassen.“ (Seite 4).
Doris Merz fand auch in der neuen Bert-Gruppe, dass das Männchen eine periphere
Stellung hat und dass die Gruppe in Untergruppen zerfällt, konzentrierte sich bei
ihren sorgfältigen methodenkritischen Beobachtungen auf den Vergleich der
Bruno-Gruppe (Saimiri sciureus) mit der Albo-Gruppe (Saimiri boliviensis). Ihre
Arbeit erbrachte: „Qualitative Unterschiede im Verhalten lassen sich nicht aufzeigen.
In beiden Sozialgruppen war zudem jeweils eines der Weibchen attraktivster
Sozialpartner. Die Weibchen suchten sich in beiden Gruppen gegenseitig häufiger auf
als das Männchen. ... Schließlich erbrachte der genaue Vergleich aber auch
eindeutige Unterschiede zwischen den beiden Varietäten des Totenkopfaffen,
besonders auffällig waren solche in der Aktivitätsrhythmik. Bei den bolivianischen
Totenkopfaffen dauert das Anfangsmaximum sechs Stunden, bei den guayanischen vier
Stunden. Nach einer einstündigen Mittagsrast sind die Individuen bis zum
Lichtwechsel aktiv, wobei das Endmaximum fünf (Bolivianer) bzw. sieben
(Guayaner) Stunden umfasst. Zudem konnten folgene Unterschiede gezeigt
werden:
Zum Rasten während der Ruheperioden suchten die bolivianischen Totenkopfaffen ein
freischwingendes Holmengerüst, die guayanischen festmontierte Ruhebretter
auf.
Das Männchen der bolivianischen Totenkopfaffen interagierte häufig mit den
Weibchen der Gruppe, das Männchen der guayanischen selten.
Bei dem Kontaktsitzen, dem Zusammensitzen in engem Körperkontakt, sitzen die
guayanischen Totenkopfaffenweibchen vornehmlich mit dem anderen Weibchen, die
bolivianischen mit dem Männchen zusammen.
Bolivianische Totenkopfaffen putzen sich häufiger und kratzen sich seltener als die
Vertreter der guayanischen Varietät.
Bolivianische Totenkopfaffen putzen einander häufig, die soziale Körperpflege
zwischen guayanischen dagegen konnte nicht beobachtet werden.“(Seiten 91 -
92).
Doris Merz hatte also unseren Eindruck, dass es sich bei den geschmuggelten Tieren
um eine andere Art handelt, durch die von ihr aufgezeigten Verhaltensunterschiede
eindeutig belegt. Dies veranlasste uns, beide Varietäten als unabhängige Species in
unserer Primatenstation zu führen.
Mit der Identifizierung des bolivianischen Totenkopfaffen als eine gesonderte Form
kamen wir zum gleichen Schluss wie R. W. Thorington in seinem Beitrag in dem
1985 erschienenen neuen „Handbook of Squirrel Monkey Research“. ([225]).
Auf seine Erkenntnisse werde ich am Ende des Kapitels noch ausführlich
eingehen.
Im Januar 1987 importierte ich dann noch weitere zehn Totenkopfaffen
(zwei Männchen (Tom und Tim), acht Weibchen) über Miami
aus British Guayana. Diese waren für die ethologische Forschung
bestimmt.
Elke Harigel begleitete diesen Import im Rahmen ihrer
Staatsexamensarbeit
vom 16.01.1987 bis zum 21.04.1987 und bebachtete die Gruppe bereits in der Quarantäne
und später im Haltungsraum. Mit ihren Beobachtungen bestätigte sie erwartungsgemäß die
bisherigen Befunde an unserer Kolonie, insbesondere die Untergruppenbildungen
innerhalb der Weibchen. Sie belegte, dass einmal gebildete Präferenzen überaus stabil
sind.
Die neue Gruppe entwickelte sich nicht zufriedenstellend, die bereits von Beginn an
vorhandenen Spannungen zwischen den Weibchenuntergruppen nahmen zu, die
Auseinandersetzungen wurden nach der erfolgreichen Geburtensaison 1987 noch
schlimmer, die Gruppe bestand aus zwei sich anfeindenden Lagern, Tim mussten wir
separat halten. 1988 verloren wir auch eines der Gründerweibchen nach einer
Bißverletzung. Zudem hatten wir weitere Saimiri - Probleme. In der Bert-Gruppe
wurden 1987 zwei neugeborene Weibchen nicht angenommen, Delhia (geb. 28.02.1987)
wurde von Elke Harigel, Dora (geb. 16.11.1987) von Birgit Zeleny mit der Flasche
großgezogen, diese mussten wir irgendwie integrieren. Nach langen Diskussionen mit
meinen wissenschaftlichen Mitarbeiterinnen, die eigentlich - wie auch ich - kein
primäres Forschungsinteresse an Totenkopfaffen hatten, wir arbeiteten bereits mit
anderen Arten und waren ausgelastet, beschlossen wir, das Problem gemeinsam zu
lösen, bzw. zu versuchen, einen Lösungsweg zu suchen. An dem Projekt beteiligt waren
neben Hella Höhmann, Barbara Jantschke, Cornelia Schäfer-Witt und mir auch eine
interessierte Studentin (Rebekka von Borstel). Das Projekt begann im Januar
1989
und war im Juli 1989 abgeschlossen. ([289])
Zuerst „sozialisierten“ wir die Handaufzuchten, wir vergesellschaften Delhia und Dora
mit dem Gründerweibchen Mara und mit Tim.
Wir stellten hierdurch zwischen diesen vier Individuen Vertrautheit experimentell her.
Die gemeinsame Haltung gelang ohne Probleme. Nach drei Monaten gemeinsamer
Haltung separierten wir die vier Individuen und ließen - begleitet von täglich
mehrstündigen parallelen Beobachtungen zu Mara, Dora und Delhia jeweils ein
Weibchen der 1987 importierten Gruppe hinzu. Nach vierzehntägiger Haltung der
Zweiergruppen, introduzierten wir ein weiteres Weibchen, wobei wir darauf achteten,
dass dieses aus dem Lager stammte, mit dem das erstintroduzierten Weibchen in der
Ausgangsgruppe nicht harmoniert hatte. Nach weiteren vierzehn Tagen fügten wir
dann noch ein weiteres Individuum hinzu (das letzte der Importweibchen und die
Männchen Tim und Tom). Wir hatten also drei Vierergruppen gebildet, die ohne
Auseinandersetzungen zusammenlebten. Im Juli schließlich fusionierten wie die
drei Gruppen, was gelang. Wir hatten erfolgreich eine große Sozialgruppe
zusammengestellt, in der Konflikte selten waren, Delhia lebte während der
Kleingruppenbildungen immer peripher und blieb es auch im neuen Verband, war aber
nicht Ziel von Aggressionen.
Tabelle 11.2: Viktorgruppe 21.09.1988 - 29.05.1993
Zusätzlich beobachteten wir gemeinsam eine am 21.09.1988
von Sigrid Hopf übernommene sechzehnköpfige Gruppe, die
Viktor-Gruppe.
Diese Gruppe, wohl kolumbianischer Totenkopfaffen, war unwahrscheinlich
wertvoll, da die Historie und die verwandtschaftlichen Beziehungen aller
Tiere durch die Arbeit von Sigrid Hopf und ihren Mitarbeitern bekannt
waren.
Ihr und dem Direktor des MPI für Psychiatrie, Detlev Ploog, der ihr die Erlaubnis
gab, ihre Tiere nach Kassel zu transferieren, möchte ich ausdrücklich für dieses
Geschenk danken. Das individuelle Kennen der Tiere war für uns wichtig. Sigrid Hopf
identifizierte in Kassel alle Individuen, und wir markierten sie anfänglich mit
Tierfarben, später mit Rasuren (besonders Kopfrasuren sind bei dem Beobachten noch
nicht individuell bekannter Tiere eine große Erleichterung). Diese Rasuren erneuerten
wir regelmäßig, bis wir sicher waren, alle Individuen zu kennen. Um das Vorwissen
nicht zu verlieren, markierten wir dann vorsichtshalber alle Tiere mit einer
Tätowierung, wobei uns hier Irmgard Küderling vom Deutschen Primatenzentrum
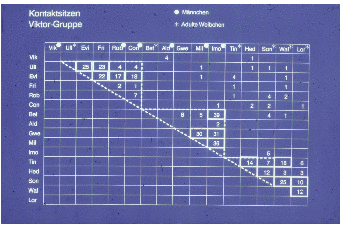
half. Nach dem Kennen aller Individuen protokollierten wir das Kontaktsitzen in der
Viktor-Gruppe (Studie W1). ([195])
Viktor hatte nahezu keine engeren Kontakte, viermal protokollierten wir das
Kontaktsitzen mit Aldo. Die Gruppe zerfiel offensichtlich in drei Untergruppen, (1)
Ulla und ihre Kinder, (2) Betty und ihre Kinder und (3) die restlichen vier adulten
Weibchen (Lore, Wally, Sonja, Tina) und Tinas Tochter Hedi.
Bereits vor der Viktorgruppe übernahmen wir von Sigrid Hopf am 06.06.1988 ihre
Nils-Gruppe und dann am 04.04.1989 ihre Karl-Gruppe. Die Tiere der letztgenannten
Gruppe waren einheitlich, sie gehörten ein- und derselben Unterart an, einer anderen
als unsere bisherigen Totenkopfaffen. Insofern berichte ich - vernachlässigen wir vorerst
Saimiri boliviensis - nun über zumindest drei Varietäten des Totenkopfaffen, (1) über
unsere Totenkopfaffen aus British Guayana, (2) über kolumbianische Totenkopfaffen
und (3) über die Individuen der Karl-Gruppe. Auch diese Gruppen beobachteten wir
regelmäßig.
Bei einer so großen Primatenkolonie ereignen sich regelmäßig auch Todesfälle. Dem
Sterben geht meist ein wochen- oder monatelanges Siechtum voraus. Tote Affen
wurden von meinen Tierpflegerinnen geborgen und in den Kühlraum gelegt. Mein
erster Gang am Morgen war dann auch immer der Weg in den Kühlraum. Tote Affen
wurden gewogen und dann in das Untersuchungsamt gebracht, wie ich es bereits in
Kapitel 8 beschrieben habe. Dieser Alltag änderte sich schlagartig am 08. November
1989 mit dem dem Tod eines Totenkopfaffen (Befund: Pseudotuberkulose). Das
Bakterium Yersinia pseudotuberculosis ist ein naher Verwandter von Yersinia pestis,
dem Erreger der Pest. Wie bei der Pest starben uns nun täglich Affen, vor allem
Totenkopfaffen. Der Krankheitsverlauf war stets derselbe. Am Abend sprangen sie
noch ohne Anzeichen von Krankheit im Gehege herum, am nächsten Morgen lagen sie
tot auf dem Boden. In dieser schrecklichen Zeit bin ich dann immer sehr früh in die
Primatenstation gefahren, um vor dem Eintreffen meiner Tierpfleger die Leichen zu
bergen, ich wollte ihnen die Belastung ersparen. Die Tiere brachten wir teilweise in
unser Veterinäruntersuchungsamt, teilweise in das Deutsche Primatenzentrum,
immer mit demselben Befund. In kürzester Zeit verloren wir so alleine dreizehn
Totenkopfaffen (neben anderen Tieren unserer Kolonie). Dr. Manfred Brack,
der Leiter der Abteilung Pathology and Veterinary Services des Deutschen
Primatenzentrums ließ einen Impfstoff passgenau für unser Bakterium herstellen
und riet uns, alle Tiere der Kolonie zu impfen. Er bot auch die personelle
Hilfe des Primatenzentrums an, seine Tierärztin Teresa Jane Gatesman wäre
bereit, die Impfung durchzuführen. Selbstverständlich nahm ich das Angebot
an.
Nun ist das Impfen von über dreihundert Affen nicht so einfach, wie eine
Massenimpfung bei uns Menschen, sie stehen nicht an, sie müssen vielmehr einzeln
eingefangen und festgehalten werden. Bei den kleineren Arten hatten wir hier gute
Routine. Bei den größeren Arten (Kapuzineraffen, vgl. Kapitel 13, und Makaken, vgl.
Kapitel 14) hingegen ist das Einfangen mit dem Netz sehr schwierig, gerade die großen
kräftigen Männchen bleiben am Gitter hängen und fliehen nicht, vielmehr halten sie
das Netz einfach fest. Bei den Makaken konnte dieses Problem vernachlässigt werden,
da sie trainiert waren, einen kleinen Zwischengang aufzusuchen (vgl. Kapitel 14). Bei
den Kapuzineraffen, die vorher nur einmal eingefangen wurden (von den kräftigen und
erfahrenen Tierpflegern des Rotterdamer Zoos (vgl. Kapitel 13), fühlte ich mich
überfordert und beschloss daher, auch die Kapuzineraffen zu trainieren, was
unwahrscheinlich schnell - viel schneller als bei den Makaken - gelang. Ich möchte hier
aber betonen, dass zwei entscheidende Unterschiede zwischen beiden Species
bestehen. Nur Makaken zeigen das Verhalten „Pressen“, sie legen sich ohne
Gegenwehr auf den Boden und leiten so die Aggressionen des Angreifers ab, nur
Kapuzineraffen waschen mit Harn, ihre Hände und Füße sind daher wie mit
Creme eingeschmiert und entgleiten daher sehr schnell der sie festhaltenden
Hand.
Am 06.01.1990 impfte Teresa J. Gatesman jeweils zwei Individuen von jeder bei uns
gehaltenene Art. Am 16.01. und 17.01.1990 erfolgte der Rest der Impfaktion.
Leider ereignete sich dabei ein Unfall, Teresa J. Gatesman wurde von einem
Kapuzineraffenmann in einen Finger gebissen, dabei wurde der Knochen getrennt.
Sofort fuhr ich mit ihr in da nächstgelegene Krankenhaus und ließ die Wunde
versorgen, anschließend impfte Frau Gatesman weiter. (Leider verlor sie den Finger
später, was mir unendlich leid tat und tut.) Nun mussten die Affen noch ein zweites
Mal geimpft werden. Ich fragte meinen Freund Dr. Walter Becker um Rat und bekam
die Antwort, er könne es machen, ich müsste nur die offizielle Genehmigung seines
Amtes einholen. Meine Bitte um Amtshilfe wurde gewährt, und gemeinsam impften
wir am 07.02. bzw. 26.02.1990 alle Individuen der Kolonie. Seit dieser Zeit
habe ich eine besondere Hochachtung vor Tierärzten. Bei beiden Impfungen
habe ich persönlich Fehler gemacht, bei der ersten Impfaktion habe nicht ich
persönlich die Beine herausgezogen, bei der zweiten Impfaktion gewährte
ich den Kapuzineraffen jeweils nach der Impfung Zugang zum Außen- und
Innengehege (in dem der Gang stand). Auch hier ereignete sich ein Unfall.
Don, das dominierende Männchen (vgl. Kapitel 13), ging ganz langsam auf
mich zu und biss mich kräftig in den Unterschenkel, was aber ohne Folgen für
mich blieb. Don hatte mich wohl als den Schuldigen enttarnt und bewies
Charakter.
Die Seuche hatte ihre Opfer in allen Totenkopfaffengruppen gefordert, mehr oder
weniger unbeschadet blieb nur unsere wertvollste Gruppe, die Viktorgruppe, hier
verloren wir nur Gwen, die Tochter von Betty. Gemeinsam mit Barbara Jantschke und
Cornelia Schäfer-Witt protokollierte ich 1992 und 1993 regelmäßig das Verhalten dieser
Gruppe (Studie W2). Die Ausgangssituation der Gruppe hatte sich geändert.
Bei den Weibchen hatten wir neben der schon erwähnten Gwen 1990 auch
Hedi (nach einer mißglückten Geburt) verloren, 1992 starben Sonja und die
uralte Lore. Die Männchen Aldo und Milo ereilte das traurige Schicksal der
Totenkopfaffennachzuchtmännchen.
Betty war nun allein, sie hatte ihre Kinder als Untergruppenpartner verloren, Gwen
starb - wie schon erwähnt - , die Söhne verließen erwartungsgemäß die mütterliche
Untergruppe. Betty fand aber Anschluss an Ulla und deren Nachkommen.
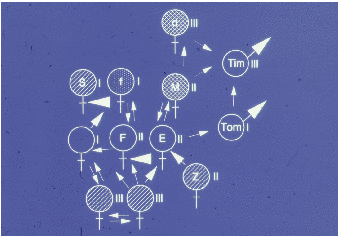
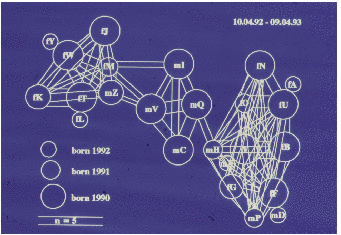
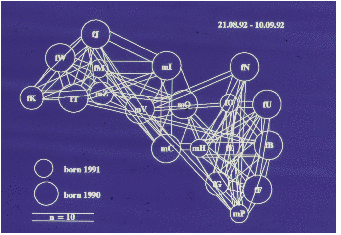
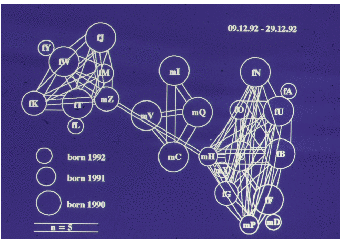
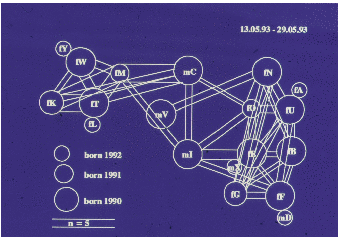
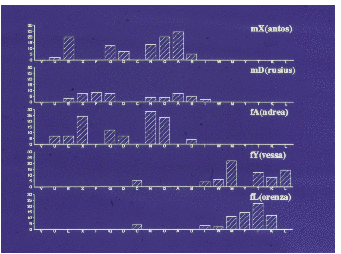
Auf der Abbildung 11.17 erkennen wir deutlich drei Untergruppen (die Werte sind
hier und auf den folgenden Abbildungen Mittelwerte pro Beobachtungsstunde), eine
Männchenuntergruppe und zwei Weibchenuntergruppen, nämlich Tina und Wally mit
ihren Töchtern und Enkelkindern bzw. Betty und Ulla gemeinsam mit Ullas Töchtern
und Enkelkindern. Vor der Geburtensaison (Abbildung 11.18) vermitteln die jungen
Männchen zwischen den Untergruppen. Nach der Geburtensaison (Abbildung 11.19)
ist die Trennung der Untergruppen noch auffälliger. In der Zuchtsaison (Abbildung
11.20) bleiben die Weibchenuntergruppen erhalten, die Männchenuntergruppe
hingegen löst sich auf. Zusätzlich haben wir die Sozialpartner der in der Gruppe 1992
geborenen fünf Jungtiere betrachtet (Abbildung 11.21), sie haben tatsächlich nur
Kontakte mit Untergruppenmitgliedern. Die scheinbare Ausnahme bei Yvessa und
Lorenza sind Kontakte zu Conny, der der mütterlichen Untergruppe nicht
angehört.
In den folgenden Jahren änderte sich nur wenig an der Konstellation der Gruppe.
Bemerkenswert ist nur die Aufzucht der Tina-Tochter Jenny. Diese wurde am Tag der
Geburt von Fritzi „gestohlen“ und von ihr aufgezogen und wuchs als Mitglied der
Ulla/Betty Untergruppe auf.
Im Rahmen ihrer Dissertation ([209]) beobachtete Pia Steinweg 1995 bis 1997 erneut diese
Gruppe..
Sie war weiter angewachsen und bestand am Ende der Untersuchung aus Individuen
von vier Generationen in Menschenobhut, eine Gruppe also, die bisher noch keinem
Totenkopfaffenforscher zur Verfügung stand.
Von den vielen Individuen der Viktorgruppe wurde die 1990 geborene Kimba meine
Freundin. Kimba steckte wohl unglücklich einen Arm durch das Gitter in eine
benachbart gehaltene Javanermakakengruppe. Dabei wurde der Unterarm
unrettbar zerbissen. Nach ausführlicher Beratung entschlossen wir uns, auf
eine völlige Amputation des Armes zu verzichten. Wir hielten stattdessen
Kimba in einem kleinen Käfig in der Futterküche und pflegten sie gesund.
Wir badeten den Arm zweimal täglich mit Kamillosan. Durch dieses Handling verlor
Kimba jegliche Scheu vor uns Menschen. Nachdem endlich der Unterarm abgefallen
war, den Oberam konnten wir erhalten, setzten wir Kimba in ihre Geburtsgruppe
zurück. Hier sprang sie sofort munter herum, ohne irgendwelche Beeinträchtigung.
Kimba hatte aber ihr Verhalten geändert. Sobald sie mich sah, sprang sie sofort
an das Gitter, um Kontakt aufzunehmen. 1994 wurde die Primatenstation
(und ich) Opfer einer Kampagne des Bundes gegen den Mißbrauch der Tiere,
was zahlreiche Fernsehteams anlockte. Immer dann, wenn Aufnahmen von
Totenkopfaffen gemacht wurden, war Kimba an vorderster Front. Machte ich die
Kameramänner auf ihr Handicap aufmerksam, wollten diese es eigentlich nicht
glauben, dass die gewandt herbeispringende Totenkopfdame eine Invalidin
war.
Tabelle 11.3: Viktorgruppe 10.08.1995 - 16.04.1997
Pia Steinweg nutzte ebenfalls den von Sigrid Hopf ([95]) erstellten Verhaltenskatalog
und analysierte in ihrer Arbeit auch nur selten zu beobachtenden Verhaltensweisen.
Zum Kontaktsitzen führt sie aus: „Die in der Gruppe geborenen Kinder beschränken
sich beim Kontaktsitzen ebenfalls auf Mitglieder der eigenen Untergruppe (und
interagieren auch spielerisch hauptsächlich mit Kindern der eigenen Untergruppe). ...
Die größere der beiden Weibchen-Untergruppen, die „Ulla-Betty“-Untergruppe, zeigt
die Tendenz zur weiteren Aufspaltung: Einige Weibchen, z.B. Fritzi, Gina und Evi,
präferieren beim Kontaktsitzen die nicht verwandte Betty vor ihrer Mutter
Ulla, so
dass durch individuelle Präferenzen Untergruppen bzw. zwei Lager innerhalb der
Untergruppe entstehen.
Für die vorgestellten Kontaktsitz-Dyaden lassen sich in den meisten Fällen
ein aktiverer bzw. ein eher passiver Interaktionspartner bestimmen. In der
Ulla-Betty-Untergruppe sind die aktiveren Initiatoren des Körperkontaktes
die jüngeren Tiere, vor allem juvenile Weibchen und Infantes. Die beiden
ältesten Weibchen Ulla und Betty nehmen aktiv insgesamt signifikant seltener
Kontakte auf (pro Individuum im Mittel 6,7 mal) als die restlichen neun
Weibchen dieser Untergruppe (im Mittel 19,6 mal) (p< 0,025). Es fällt aber auf,
dass sie im Vergleich zu den anderen Untergruppenmitgliedern am häufigsten
aufgesucht werden (17,34 mal gegenüber 9,04 mal bei den sechs anderen
Weibchen)
Die beiden Gründerweibchen sind also für adulte bzw. fast adulte Weibchen die
attraktivsten Kuschelpartner. ... In Mutter-Tochter-Dyaden - die alten Weibchen und
auch die Dyade Fritzi-Jenny (s. o.) eingeschlossen - ist die Tochter in der Regel der
aktivere Partner. Auch für die verschwisterten Weibchen verschiedenen Alters gilt,
dass zumeist das jüngere Tier häufiger den Kontakt initiiert. Einzige Ausnahme bildet
die Dyade Ulla mit ihrer ein Kind tragenden Tochter Evi, zu der sie jedoch nur
während der Aufzuchtsphase affiliativen Kontakt aufnimmt. Kinder tragende
Weibchen sind für andere Tiere generell attraktiver als während des restlichen
Zyklus.
Das Kontaktsitzen zwischen Männchen und Weibchen kommt nur vernachlässigbar
selten und ausschließlich während der Paarungszeit vor. ... Wie bereits erwähnt,
kommt das Kontaktsitzen zwischen den Individuen der verschiedenen Untergruppen
nicht vor; es gibt keine einzige Abweichung, ... “ ([209], Seiten 44 - 46).
Während der Beobachtungen von Steinweg war allein Imo reproduktiv aktiv. Nach
Steinweg ([209]) ist die olfaktorische Kontrolle der Genitalien durch eines der
Männchen das auffälligste Merkmal der Paarungszeit. In der Viktorgruppe konnte
Steinweg auch soziale Körperpflege beobachten, im Wochenmittel beobachtete sie
dieses Verhalten 0,03 mal pro Tier. Insgesamt protokollierte sie 52 Putz-Dyaden,
51 gingen von Weibchen aus, einmal putzte Imo ein Weibchen während der
Paarungszeit.
Agonistisches Verhalten beobachtete Steinweg in der Viktorgruppe
im Wochenmittel pro Tier 0,028 mal. Vor allem beobachtete sie das
„Jagen“.
Innerhalb der Weibchen war das Jagen nur von Individuen der Betty/Ulla-Untergruppe
gegenüber Mitgliedern der Tina-Wally-Untergruppe zu beobachten. Individuen beider
Weibchenuntergruppen jagten die drei Männchen, vor allem während der Paarungs-
und Geburtenphase. Besonders häufig wurde Conny gejagt (von seiner Mutter Ulla,
seiner Schwester Evi und von der Tina-Tochter Lorenza).
Das genitale Imponieren (s. o.) wurde ebenfalls von Steinweg protokolliert und
ausgewertet, es trat in der Viktorgruppe im Wochenmittel 0,016 mal pro Tier auf,
juvenile Weibchen und Kinder zeigten dieses Verhalten häufiger als adulte Individuen.
Es wurde auch zwischen den Individuen ein- und derselben Untergruppe beobachtet,
eine Beziehung zu positiv-sozialem oder agonistischem Verhalten konnte nicht
aufgezeigt werden.
Pia Steinweg beobachtete auch die Bruno-Gruppe (05.07.1995 - 07.09.1996), eine von
uns nach der Seuche neugebildete Gruppe von Totenkopfaffen aus British
Guayana. Die Gruppe bestand aus dem schon bekannten Bruno, einem der
1987 importierten Wildfangweibchen, deren Tochter und einem weiteren vor
der Seuche geborenem Nachzuchtweibchen des 1987-Importes und Kindern
dieser drei Weibchen. Auch in dieser kleinen Gruppe konnten eindeutig zwei
Untergruppen aufgezeigt werden, beruhend auf Verwandtschaft. Bemerkenswert ist
auch, dass Pia Steinweg nur dreimal soziale Körperpflege beobachten konnte,
wobei jeweils ein Kind von seiner Mutter bzw. Großmutter geputzt wurde.
Bruno mussten wir am 22.10.1995 (20 Jahre nach Import) aus der Gruppe
entfernen, da sein Allgemeinzustand ein Verbleiben im Sozialverband nicht mehr
gestattete.
Vom 06.07.1995 - 16.04.1997 erhob Pia Steinweg auch Daten der Karl-Gruppe.
Eigentlich reihten sich ihre Beobachtungen in die der anderen Gruppen ein, sie
belegte zwei Weibchenuntergruppen, beruhend auf Verwandtschaft, und eine
Männchenuntergruppe. Reproduktiv aktiv war während ihrer Beobachtungen vor
allem Piet, aber auch Karl zeigte Interesse an einem der alten Weibchen. Sie
beobachtete auch in dieser Gruppe soziale Körperpflege. Eindeutig einer Art/Unterart
zuordnen konnte ich diese Totenkopfaffen nicht. Diese Gruppe ist für mich rätselhaft
geblieben. Ich gebe daher in der Tabelle 11 alle mir vorliegenden Informationen.
Tabelle 11.4: Didi-Gruppe 17.03.1983 - 16.04.1997
1983 bildete Sigrid Hopf aus einem Männchen (Didi) und vier Weibchen, die einer
1978 importierten Sendung entstammten, eine neue Gruppe, die Didi-Gruppe. Drei
dieser vier Weibchen reproduzierten erfolgreich, die Gruppe wuchs - vergleichbar den
bereits besprochenen Gruppen - sukzessive an. sie bestand Ende 1987 aus elf
Individuen, den Gründertieren und sechs Nachzuchten (vier Männchen und zwei
Weibchen). 1988 starb dann Didi plötzlich. Die drei 1988 geborenen Jungtiere (ein
Männchen, zwei Weibchen) wurden aller Wahrscheinlichkeit nach (Alter der
Nachzuchtmännchen, Tragzeitlänge etwa fünf Monate) noch von ihm gezeugt. Mit dem
Tod von Didi war diese Gruppe praktisch „tot“. Die „Last“der Nachzuchtarbeit
mussten nun die in der Gruppe geborenen Männchen leisten. 1989 gab Sigrid Hopf
diese Gruppe an uns ab. Hier ereilte auch diese Gruppe die oben beschriebene Seuche,
wir verloren die nicht reproduktive Fanny und ein Nachzuchtmännchen. Ab 1990
züchtete auch diese Gruppe, ab 1991 auch in zweiter Generation. Für mich völlig
unerklärbar ist die hohe Überlebensrate der Nachzuchtmännchen, 1995 starb
zwar ein fast achtjähriges Männchen (Tommy), doch war auch dieses Tier
unverletzt.
Helmi entnahm ich 1996 zehnjährig für eine Untersuchung, über die ich in Kapitel 15
berichten werde. Möglichweise könnte für das Überleben die geringe Anzahl der
Weibchen verantwortlich gewesen sein (s. u.), auszuschließen ist aber auch nicht, dass
hier ein Unterschied im Verhalten zwischen den Varietäten belegt worden ist (s.
u.).
Parallel zu den Beobachtungen an Saimiri sciureus war es mein Ziel, auch Saimiri
boliviensis bei uns beobachten lassen zu können, ich wollte gerne die von Doris Merz
erhobenen Befunde (s. o.) an weiteren Individuen überprüfen. Das bereits
erwähnte Trio (Albo/Rika/Gesa) zeigte keinerlei reproduktive Aktivitäten,
offensichtlich war diese Gruppe zu klein und/oder zu vertraut. Nicht wissend, was in
naher Zukunft auf mich zukommen sollte, kaufte ich 1991 die bolivianischen
Totenkopfaffengruppe von Lord Fisher aus Kilverstone, ein Männchen (Pascal) und zehn
Weibchen
unbekannten Alters, die Geburtsdaten, die ich in der Fußnote in Klammern angegeben
habe, sind von uns geschätzt; diese importierten wir am 29.01.1992 und sie lebten sich gut bei
uns ein.
Reproduktive Aktivitäten konnten wir nicht beobachten. Insofern hatten wir nun zwei
Gruppen nicht züchtender Totenkopfaffen. Am 23.11.1992 starb Pascal. Ich weiß nicht,
wie alt er war, sein Gewicht (1250 g) spricht dagegen, dass er am Stress gestorben
ist, er hatte auch keine sichtbaren Verletzungen. Wir brachten ihn in das
Veterinäruntersuchungsamt. Der Bericht liegt mir nicht mehr vor, wahrscheinlich
ist der aktuelle Ordner mit den Veterinärbefunden - wie auch die Affen der
Primatenstation - „verloren“ gegangen.
Wir fusionierten daraufhin unsere beiden Bolivianer-Gruppen zur Albo-Gruppe. Dies
gelang. Der seit 1983 bei uns unreproduktiv lebende Albo schwängerte sieben der zehn
Kilverstoner-Weibchen, drei (Ickwa, Wendy und Blacki) trugen die Jungen aus, wobei
nur Blacki ihr weibliches Junges (Tonia) annahm. Die beiden anderen Jungtiere
versuchte Ursula Schäfer per Hand aufzuziehen, was bei der Ickwa-Tochter (Quicka)
gelang, die Handaufzucht des kleinen Männchens scheiterte, es starb einen
Tag nach der Geburt. Dennoch empfanden wir dieses Zwischenergebnis als
glücklichen Beginn einer erfolgreichen Zukunft. Mit Ausnahme der offensichtlich
uralten Jolanthe hatten alle adulten Weibchen ihre Zuchtfähigkeit unter Beweis
gestellt.
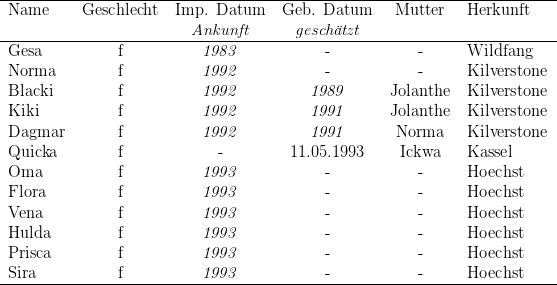
Am 22.07.1993 übernahm ich dann noch eine achtköpfige nichtreproduktive Gruppe der
Hoechst-AG,
wir gaben den Totenkopfaffen die Namen Magnum, Apollo, Oma, Flora, Vena, Hulda,
Prisca und Sira. Magnum starb am 10.09.1993 an einer Gongylonema-Infektion (vgl.
Kapitel 8). Die übrigen Tiere schienen zu gedeihen.
Im Oktober 1993 starben dann Albo, Rika, Ickwa und Tonia. Bei den ersten drei
Tieren konnten wir nicht ausschließen, dass sie den normalen Alterstod gestorben
sind,
doch vermuteten wir (nach meinen Aufzeichnungen), dass ursächlich für deren Tod
soziale Auseinandersetzungen gewesen sind; wahrscheinlich war auch Gesa
Adressat von Attacken durch Kilverstoner-Weibchen, doch wurde dies nicht
protokolliert.
Im Dezember 1993 versuchten wir dann, die ehemalige Albo-Gruppe
(Kilverstoner-Weibchen und den Wildfang Gesa), die handaufgezogene Quicka und die
von Hoechst erhaltene Gruppe (Apollo-Gruppe) zu fusionieren, dies misslang.
Die Kilverstoner-Weibchen griffen die Hoechster-Weibchen massiv an. Wir
separierten daher die Hauptaggressorinnen (Wendy, Cindy, Jolanthe, Leni
und Endla), danach fanden keine agonistischen Auseinandersetzungen mehr
statt. Eines der älteren Kilverstoner-Weibchen (Norma), drei der jüngeren
Kilverstoner-Weibchen (Blacki, Kiki, Dagmar), die handaufgezogene Quicka und
unsere alte Dame Gesa waren durch dieses Experiment relativ friedlich mit den
Hoechster-Weibchen vereint. Die übrigen Kilverstoner-Weibchen hielten wir
separat.
Am 16.02.1994 tauschte ich dann noch (gegen eine Kapuzineraffengruppe) vier
bolivianische Totenkopfaffen des Zoos in Hof ein. Diese Gruppe züchtete nicht und
bestand aus einem Männchen (Tito), zwei Weibchen (Ronja und Yvonne) und einem
Kastraten (Xaver). In diese Gruppe integrierten wir bereits einen Tag nach Ankunft in
Kassel drei der Kilverstoner Weibchen (Wendy, Endla und Jolanthe). Als dies gelang,
introduzierten wir auch Leni und Cindy in die Tito-Gruppe. Auch diesmal belegte das
neue Männchen seine Zeugungsfähigkeit, er schwängerte Cindy und Wendy, Cindy
abortierte, Wendys am 13.09.1994 geborene Tochter Ursula wuchs in der Gruppe
heran.
1995 wurden Cindy, Wendy und Leni in der Tito-Gruppe schwanger und zogen ihre
Jungen (Juno, Anna und Ingo) auch auf, wir hatten in dieser Gruppe nun mehrere
potentielle Mutter-Kind-Untergruppen (Leni, Endla, Ingo; Cindy-Juno; Wendy,
Ursula, Anna).
Am 01.04.1995 starb dann Apollo, die Apollo-Gruppe war ohne Männchen. Das
Todesgewicht von Apollo (890 g) erlaubt die Spekulation, dass auch Apollo Opfer der
Weibchen seiner Gruppe geworden ist. Das Risiko, nun beide Gruppen erneut
zu fusionieren, wollten wir so nicht eingehen. Nach reiflicher Beratung mit
meinen Mitarbeitern hielt ich es nur für vertretbar, die Totenkopfaffen mit einer
Männchengruppe zu fusionieren, in der Hoffnung, mehrere Männchen könnten
dem Druck der Weibchen standhalten. Zudem hielt ich es für wichtig, alle
Versuchsvorhaben begleitend zu beobachten, hier war meine Doktorandin Pia Steinweg
eine nicht ersetzbare Hilfe.
Ich möchte nicht verschweigen, dass ich vor der Bildung der Gruppe gewarnt worden
bin, man könne nicht mehrere erwachsene Männchen gemeinsam halten. Aus einem
deutschen Zoo hörte ich, dass sich die Männchen bolivianischer Totenkopfaffen
gegenseitig sukzessive umgebracht hätten (jeweils wäre diesen Todesfällen ein
Tumult vorausgegangen, die Handelnden hätte man nicht identifizieren können).
Heute bin ich sicher, diese „Fakten“ beruhen auf Nichtbeobachten und auf der
Annahme, dass Affenmännchen untereinander intolerant sind. Nach meinem
heutigen Wissen bin ich überzeugt, dass nicht die Männchen sich gegenseitig
umgebracht haben, vielmehr dürften sie sukzessive Opfer der Weibchen geworden
sein.
Am 19.08.1995 übernahmen wir von einem privaten Halter den im März 1986 geborenen Hans
und den am 15.07.92 geborenen Muck, beide Tiere kannten sich nicht. Zudem holte ich 1996 aus
Apenheul
die gemeinsam gehaltenen adulten Männchen Jansen (geb. 1986) und Youp (geb. 1990)
und ein 1992 geborenes jungadultes Männchen (Yatzee), der den beiden anderen
Männchen unvertraut war.
Die mit ihr und meinen Mitarbeitern ausführlich diskutierte Aufgabe von Pia
Steinweg war es nun, nicht nur die beiden bestehenden Sozialgruppen zu beobachten,
sie sollte alle Weibchen unter Beobachtung fusionieren, unsere Männchen
zusammenzuführen und anschließend die Männchengruppe integrieren. Ihre
Dissertationsarbeit war anspruchsvoll.
Tabelle 11.5: Tito-Gruppe 1995
Pia Steinweg beobachtete die Tito-Gruppe vom 01.08.1995 bis zum 18.01.1996.
Bedrohliche agonistische Interaktionen konnte sie in dieser Zeit nicht feststellen, doch
war in diesem Sozialverband auffällig, dass Empfänger agonistischer Interaktionen
ausschließlich die beiden Männchen waren. Vor allem Xaver wurde von Wendy, Ronja,
Ursula und Leni bedrängt. Eine deutliche Untergruppenbildung war nicht erkennbar.
„Enge Kontakte bestehen zwischen verwandten Tieren (Leni-Endla-Ingo;
Wendy-Ursula-Anna; Cindy-Juno), jedoch halten alle Tiere untereinander Kontakte. In
den Dyaden, an denen die Infantes beteiligt sind, sind diese in der Regel der aktivere
Partner bei der Kontaktaufnahme. Aus der Vorgeschichte der Tiere ist bekannt, dass
die beiden Weibchen Ronja und Yvonne länger miteinander vertraut sind als mit den
restlichen Weibchen. ... sie (halten) jedoch mit anderen Weibchen mehr Kontakte als
untereinander. ... Ronja und Yvonne sind also in die Gruppe der Weibchen
Kilverstoner Herkunft integriert (was sich auch an ihren engen Kontakten zu
den Kindern zeigt, bei denen sie sich maßgeblich an der Fremdelternpflege
beteiligen).
Der auffälligste Unterschied zu S. sciureus liegt im Kontaktsitzen zwischen
den Geschlechtern, das in dieser Gruppe unabhängig von der Paarungssaison
häufig zu beobachten ist. Somit gibt es hier weder eindeutige Weibchen- noch
Männchen-Untergruppen. Die gemeinsame Vorgeschichte der beiden Männchen und
der Weibchen Ronja und Yvonne scheint nur bedingt Einfluß auf ihre gegenseitigen
affiliativen Beziehungen zu haben: Während der reproduktiv aktive Tito keinen
Kontakt zu ihnen hält, besteht zwischen dem Kastraten Xaver und Yvonne eine realtiv
enge Bindung ..., wobei Xaver der aktivere Partner bei der Kontaktaufnahme ist.
... Von den adulten Weibchen initiieren die (kinderlosen) Weibchen Endla,
Ronja und Yvonne signifikant häufiger den Kontakt als die drei Weibchen mit
Nachkommen. ... Die Kinder und das juvenile Weibchen sind auch in diesem
Sozialverband am aktivsten bzgl. der Kontaktaufnahme.“ ([209], Seiten 56 -
59).
Weitaus häufiger als in den von ihr beobachteten drei Saimiri sciureus-Gruppen
protokollierte Steinweg soziale Körperpflege, Empfänger waren vor allem das
juvenile Weibchen Ursula und die drei Kinder. Gegenseitige soziale Körperpflege
beobachtete sie nur zwischen Leni und Endla, die wir (s. o.) als Lenis Tochter
identifiziert hatten. Die beiden ältesten Weibchen (Leni und Wendla), beteiligten
sich kaum an der sozialen Körperpflege, sie werden aber von den jüngeren
Weibchen Endla und (seltener) Cindy geputzt, also von Weibchen gleicher
(Kilverstoner) Herkunft. Den kastrierten Xaver putzen ebenfalls diese zwei jungadulten
Weibchen und das Hofer-Weibchen Yvonne. Yvonne ist zudem das einzige
Weibchen, das regelmäßig Tito putzt; weit häufiger putzt sie aber Ursula
und Juno. Die Männchen nahmen aktiv nicht an der sozialen Körperpflege
teil.
Auch das Genitale Imponieren beobachtete Steinweg in dieser Gruppe häufiger als bei
Saimiri sciureus, dies gilt für beide Geschlechter. Häufigster Empfänger dieses
Verhaltens war Leni, die mit Ausnahme ihrer Tochter Endla von allen adulten
Weibchen „animponiert“ wurde, vor allem von Cindy. Tito zeigte dieses Verhalten
gegenüber Endla und Cindy, den von ihm präferierten Kilverstoner Weibchen.
Bemerkenswert ist vielleicht noch, dass der kleine Juno gegenüber Xaver relativ häufig
imponierte.
Tabelle 11.6: Nur-Weibchen-Gruppe 1995
Pia Steinweg beobachtete die Nur-Weibchen-Gruppe vom 05.10.1995 bis zum
18.01.1996. Das Zusammenleben war friedlich, nur einmal musste Steinweg das Jagen
protokollieren (Flora jagt Vena). In dieser Gruppe war eine Untergruppe
aufzeigbar, so hielten fast alle der Hoechster Weibchen untereinander engen
Kontakt, an diese Untergruppe konnte sich die 1983 importierte Gesa
anschließen.
Das wohl älteste Weibchen der Gruppe Oma hält keine Kontakte zu anderen
Gruppenmitgliedern. Geringere Kontakte beobachtete Steinweg zwischen der
Mutter-Tochter-Dyade Norma/Dagmar, die Kontakte gingen von Dagmar aus; das
nach Norma älteste Kilverstoner Weibchen Blacki initiiert keine Kontakte, wird aber
von seiner jüngeren Schwester Kiki und der handaufgezogenen Quicka aufgesucht.
Der einzige enge Kontakt zwischen einem Hoechster Weibchen (Prisca) und
einem Kilverstoner Weibchen (Norma) geht von Prisca aus. (Der seltenere
Kontakt der Norma-Tochter Dagmar zu Prisca hingegen wird von Dagmar
initiiert.) Auffällig in dieser Gruppe war wiederum das im Vergleich zu Saimiri
sciureus häufige soziale Putzen, vor allem durch die jüngeren Kilverstoner
Weibchen. Gegenseitige soziale Körperpflege wird nur zwischen den Schwestern
Blacki und Kiki beobachtet. Am häufigsten wird das Putzen zwischen Norma
und ihrer Tochter Dagmar protokolliert, wobei hier nur Norma aktiv putzt.
Das Genitale Imponieren konnte Steinweg auch in der Nur-Weibchen-Gruppe belegen,
es ging stets von einem Partner der entsprechenden Dyade aus, selbst Oma zeigte
dieses Verhalten aktiv (gegenüber Vena und Prisca, also gegenüber Weibchen
identischer (Hoechster)-Herkunft).
Am 09.01.1996 separierten wir Tito und Xaver aus der Tito-Gruppe. Pia Steinweg
beobachtete die Restgruppe und fand keine auffälligen Veränderungen im
Verhalten der Individuen. Am 18.01.1996 erfolgte die Fusion beider Gruppen
(Nur-Weibchen-Gruppe und die restlichen Individuen der Tito-Gruppe). „Um dies zu
ermöglichen, muss lediglich ein trennendes Gitter entfernt werden, da die Tiere schon
seit Beginn der Studie Gitterkontakt, d. h. visuellen, olfaktorischen und akustischen
Kontakt haben. Zunächst herrscht einige Minuten lang auffällige Stille, wobei alle
Tiere den gesamten Käfig inspizieren und sich vermischen. Dann finden sich die
Gruppen in ihrer alten Zusammensetzung und in den jeweils vertrauten Käfigteilen
wieder ein. Kurz darauf folgen erste agonistische Interaktionen zwischen beiden
Gruppen, wobei die beiden Hofer Weibchen Ronja und Yvonne das Angriffsziel
bilden. Diese Weibchen sind als einzige Individuen für die (Mitglieder der
Nur-Weibchen-Gruppe) fremd. Die Aggressionen gehen von mehreren Tieren der
(Nur-Weibchen-Gruppe) aus. Auch andere Weibchen der (ehemaligen Tito-Gruppe)
werden - jedoch weniger vehement - angegriffen. Aggressoren sind die Hoechster
Weibchen, die bei einer Fusion im Dezember 1993 angegriffen worden waren, und zwar
von denjenigen Kilverstoner-Weibchen, die nun ebenfalls Ziel der aggressiven Übergriffe
sind. In den folgenden Tagen finden immer wieder Angriffe auf Ronja und Yvonne
statt, so dass sie am 24.01.1996, also sechs Tage nach der Fusion, aus der Gruppe
genommen werden, ... Die restlichen Weibchen behalten auch bis zur endgültigen
Fusion mit der Männchen-Gruppe am 20.03.1996 ihre räumliche Trennung
bei, wobei die (Nur-Weibchen)-Untergruppe (die Mitglieder der ehemaligen
Nur-Weibchen-Gruppe) den deutlich größeren Teil des Geheges einnimmt. Agonistische
Interaktionen werden jedoch bis dahin nicht mehr beobachtet.“([209], Seiten 152 -
153).
Die fusionierte Gruppe beobachtete Pia Steinweg drei Monate lang.
Kontaktsitzen zwischen den Mitgliedern beider Untergruppen kommt praktisch
nicht vor (abgesehen von einem einmaligen Kontakt zwischen Wendy und
Gesa). Nach den Kontaktsitzdyaden zerfielen die beiden Untergruppen
(Nur-Weibchen-Untergruppe und Rest-Tito-Untergruppe) in weitere Untergruppen. In
der Nur-Weibchen-Untergruppe hielten (1) die Hoechster-Weibchen und Gesa
engen Kontakt, (2) Norma und Dagmar und (3) Blacki, Kiki und Quicka,
wobei der Kontakt der Mutter-Tochter Dyade (Norma/Dagmar) besonders eng
war.
Die Rest-Tito-Untergruppe untergliederte sich nach Verwandtschaft: (1) Wendy und
ihre Kinder, (2) Leni und ihre Kinder und (3) Cindy und ihr Sohn Juno; engere
Kontakte innerhalb der adulten Weibchen dieser Untergruppe waren nur zwischen
Wendy und Endla beobachtbar, die Kinder vermittelten zwischen den Weibchen dieser
Untergruppe. Soziale Körperpflege beobachtete Steinweg nur innerhalb der
Untergruppen, nämliches galt für das Genitale Imponieren.
Parallel zu den Beobachtungen der Weibchen bildete Pia Steinweg sukzessive die
geplante Männchengruppe. In ihrer Arbeit ([209]) dokumentierte sie ausführlich die
einzelnen Versuchsabschnitte und berichtet über ihre Beobachtungen. Hier sollen diese
Ergebnisse nur sehr verkürzt wiedergegeben werden. Am 29.08.1995 fusionierte sie das
adulte Männchen Hans mit dem juvenilen Männchen Muck. Nach einer Phase
des Kennenlernens wird in den vier Monaten der gemeinsamen Haltung kein
agonistisches Verhalten protokolliert. Häufig hielten diese beiden Männchen
Körperkontakt, die Kontaktaufnahme ging in der Regel von Muck aus. Nach der
Separation aus ihrer Gruppe am 09.01.1996 werden Tito und Xaver in einen
Käfig mit Gitterkontakt zu Hans und Muck verbracht. „Zwei Tage später
zeigen die beiden kräftigeren Männchen der beiden Gruppen (Hans und Tito)
auffällig viel Vokalisation bei fast synchronem Verhalten; beide halten sich zur
selben Zeit am Boden auf; als Tito hochklettert, folgt Hans auf der anderen
Gitterseite. Unter beiderseitiger Vokalisation nähern sie sich einander, bis sie
schließlich Körperkontakt am Gitter haben. ... Weitere fünf Tage später haben
Tito und Hans erneut Körperkontakt am Gitter. ... (Das) trennende Gitter
(wird) nun entfernt.“ ([209], Seite 154). Nach anfänglichen Aggressionen von
Xaver gegen Muck, verläuft auch diese Fusion friedlich. Bereits am folgenden
Tag suchen sich Hans und Tito gegenseitig zum Kontaktsitzen auf. In den
folgenden zwei Monaten konnten keine agonistischen Interaktionen beobachtet
werden.
Am 21.02.1996 holte ich die drei Männchen (Jansen, Youp, Yatzee) aus Apenheul,
bereits einen Tag später fusionierte Steinweg Yatzee mit den bereits vorher
gemeinsam gehaltenen adulten Männchen. „Unter den beiden vertrauten
adulten Männchen wird soziale Körperpflege beobachtet. Auch in diesem
Sozialverband nimmt überwiegend das jüngere Männchen den Kontakt zu den beiden
adulten Tieren auf, während diese sich gegenseitig aufsuchen.“([209], Seite
156). Am 05.03.1996 werden die drei Männchen in das der Vierer-Gruppe
benachbarte Gehege umgesetzt. „(Am 06.03.1996) zeigt Hans ein recht auffälliges
Verhalten, indem er von Zeit zu Zeit unter lautstarker Vokalisation an das
trennende Gitter springt bzw. daran rüttelt. ... Innerhalb der Dreiergruppe
überwiegt während der Beobachtungszeit das Kontaktsitzen, wobei Jansen der
beliebteste Partner ist. ... Da die Tiere vier Tage später keine auffälligen
Reaktionen aufeinander zeigen, wird das trennende Gitter am 11.03.1996
entfernt.“ ([209], Seite 156). Hans jagt und verfolgt die neuen Männchen. Am
12.03.1996 werden Hans und Tito mit Bißwunden vorgefunden. Hans wirkt
eingeschüchtert, Tito hingegen macht einen kräftigeren Eindruck, imponiert
gegenüber den anderen Männchen. Bereits am 13.03.1996 sitzen Youp und
Tito im Körperkontakt. Am 14.03.1996 wird kein agonistisches Verhalten
beobachtet.
Insofern schien es vertretbar zu sein, die beiden allein gehaltenen Hofer-Weibchen
(Ronja und Yvonne) in diese Männchen-Gruppe zu introduzieren. Xaver nimmt sofort
Kontakt zu diesen ihm vertrauten Weibchen auf. „Etwa eine halbe Stunde nach
Beginn der Fusion verfolgt eines der Weibchen, Yvonne, das juvenile Männchen Muck
und später auch den subadulten Yatzee. ... Am folgenden Tag (15.03.1996) wird das
juvenile Männchen von beiden Weibchen gejagt; ... Während eines Angriffs Yvonnes
auf Muck kommt Hans hinzu, hält Yvonne fest und zieht an ihr, so dass der Eindruck
entsteht, dass er Muck unterstützt. Nach einem weiteren Tag erscheint die Gruppe
ruhig, und es werden keine agonistischen Interaktionen beobachtet. Tito sitzt nun
mit den beiden „fremden“ Männchen Jansen und Youp Kontakt.“ ([209],
Seiten 158 - 159). Vor der für den 20.03.1996 geplanten Gesamtfusion wird
(unbeobachtet) Hans erheblich verletzt, so dass er vor der Fusion separiert werden
muss.
„Die Männchen-Gruppe (nebst Ronja und Yvonne) wird zunächst in das der
Weibchen-Gruppe benachbarte Gehege gebracht, worauf kurzer Gitterkontakt zwischen
einigen Individuen beobachtet wird. Die Vokalisation und die Erregung der
Tiere nimmt stark zu. ... Nach dem Entfernen des trennenden Gitters beginnt
zunächst die bei allen Fusionen übliche Vermischung aller Tiere, wobei die jeweils
„neue“ Käfighälfte inspiziert wird. Die beiden männlichen Jungtiere (Juno und
Ingo) werden von ihren Müttern (Cindy und Leni) getragen, was in diesem
Alter eigentlich nicht mehr üblich ist. Schon bald findet ein erster Angriff
der Hoechster Weibchen auf die beiden Hofer Weibchen Ronja und Yvonne
statt, ... Außerdem wird das Hofer Männchen Xaver von ihnen angegriffen,
ebenso wie die Männchen Youp und Tito. Die introduzierte Gruppe bildet in
einer Ecke des Geheges einen dichten Pulk, der einen Angriff auf einzelne
Tiere verhindert, bis eines der Tiere sich daraus löst. Innerhalb der nächsten
Stunden werden die drei Hofer Tiere immer wieder angegriffen, wobei sich
eine Hauptaggressorin erkennen lässt. Dieses Weibchen, Sira, wird von der
Gruppe separiert. Die drei adulten Männchen sind bald für einige Weibchen
attraktiv, zu ihnen wird positiver Sozialkontakt aufgenommen. Auch nach
der Fusion nehmen die Weibchen der ehemaligen Tito-Gruppe weniger als
ein Drittel des Geheges ein, während die ehemalige (Nur-Weibchen)-Gruppe
sich im restlichen Raum verteilt. Die neu introduzierten Männchen bleiben
peripher. Diese räumliche Aufteilung bleibt in den kommenden sieben Monaten
bestehen.
Am (Tag nach der Fusion) wird das Hofer Männchen Xaver mit einer Kopfverletzung
aufgefunden, ... so dass er abends separiert wird. Daraufhin wird das juvenile
Männchen Muck von mehreren Weibchen der ehemaligen (Nur-Weibchen)-Gruppe
angegriffen, worauf er meist zu den Weibchen der (ehemaligen) Tito-Gruppe flüchtet.
Zwischen den beiden Hofer Weibchen (Ronja und Yvonne) und den (ihnen vertrauten)
Kilverstoner Weibchen ... werden erste Annäherungen beobachtet, sie halten sich
jedoch noch überwiegend in der Nähe der Männchen auf. Nach zwei Tagen sind sie
jedoch wieder räumlich in die (ehemalige) Tito-Gruppe integriert. Zwei der adulten
Männchen (Jansen und Youp) sind weiterhin attraktiv für einige Weibchen, was sich
an der häufigen Kontaktaufnahme zeigt. Am folgenden Tag (22.03.1996) weist das
Weibchen Ronja neue Verletzungen auf. Außer dem juvenilen Muck wird nun auch das
subadulte Männchen Yatzee verfolgt. Eine Unterstützung der adulten Männchen bleibt
aus. Die beiden jungen Männchen werden am nächsten Tag erstmals kontaktsitzend
beobachtet. Da sie Ziel der Aggressionen bleiben, und viele Weibchen sich an den
Verfolgungen beteiligen - darunter auch Weibchen der ehemaligen Tito-Gruppe -,
werden die beiden jungen Männchen fünf Tage nach der Fusion separiert. ...
Bei den am folgenden Tag beobachteten agonistischen Übergriffen auf Tito
wird auch dieser nicht von den anderen Männchen unterstützt. Da sich ein
weiteres Weibchen, Hulda, als Hauptaggressorin erweist, wird es ebenfalls
separiert. Dennoch wird Tito in den folgenden Tagen weiterhin von den Weibchen
der ehemaligen (Nur-Weibchen)-Gruppe agonistisch verfolgt, auch die Hofer
Weibchen Ronja und Yvonne sind mitunter noch Ziel der agonistischen Übergriffe.
...
Sechs Tage nach der Fusion sind erste reproduktive Aktivitäten zwischen Jansen und
der ihm eine Woche länger vertrauten Yvonne zu beobachten. In den folgenden
Wochen sind sowohl er als auch Youp häufige Sexualpartner der Weibchen.
Angriffe auf Tito werden in den nächsten Wochen zwar noch beobachtet.
Diese werden aber stetig seltener und weniger vehement. Nach zwei Wochen erscheinen
Ronja und Yvonne deutlich erholt, sind aber noch immer sehr vorsichtig beim
Annähern eines der fremden Weibchen. In den kommenden Monaten finden noch
einzelne Angriffe auf die drei Hofer Tiere Ronja, Yvonne und Tito statt, ... Etwa fünf
Wochen nach der Fusion (Anfang Mai) erscheint auch Tito physisch wieder völlig
gesund.
Nach Abschluss der Paarungsphase werden auch die kräftigsten adulten Männchen
(Jansen und Youp) von den Weibchen angegriffen: Mitte Mai wird erstmals
beobachtet, dass das zweitkräftigste Männchen Youp Ziel aggressiver Handlungen der
Weibchen ist, zwei Wochen später auch das kräftigste Männchen Jansen.“ ([209],
Seiten 159 - 161).
Hier endet „mein“ Bericht über die bolivianischen Totenkopfaffen, ab Mitte Juni 1996
(vgl. Kapitel 15) mussten die Totenkopfaffen ohne meine mehrfachen täglichen
Beobachtungsgänge auskommen. Ich konnte nur noch Pia Steinweg persönlich bzw.
meine verbliebenen Mitarbeiter telefonisch beraten. Es steht mir auch nicht zu,
irgendwelche Entscheidungen zu kritisieren, da ich selber über Monate abwesend und
persönlich nicht verfügbar war.
“Gelegentlich wird auch noch ein weiteres Weibchen der ehemaligen Tito-Gruppe,
Cindy, verfolgt, jedoch nicht angegriffen. Dieses Weibchen wird Ende Juni
dabei beobachtet, wie es auf einen Angriff auf Ronja und Yvonne mit einem
Gegenangriff reagiert. Das angreifende Weibchen weicht vor ihr. Cindy scheint
etwa zu diesem Zeitpunkt die Untergruppe zu wechseln.“ ([209], Seiten 161 -
162).
Die Monate Juli bis September 1996 waren ereignisreich, Mitte August muss Tito, Ende
August Youp (jeweils nach heftigen Angriffen der Weibchen) aus der Gruppe entfernt
werden. Am 23.07.1996 stirbt Vena. Von den Weibchen der Tito-Untergruppe ist zu
berichten, dass Wendy am 05.07.1996 abortierte, die bisher kinderlose Endla hatte am
14.08.1996 einen Spätabort. Innerhalb der Nur-Weibchen-Untergruppe trägt die bisher
(13 Jahre lang in Kassel) kinderlose Gesa ihr Jungen aus, dieses ist aber wohl
übertragen (Geburtsgewicht: 100 g) und wird am 23.08.1996 tot vorgefunden, Cindys
Jungtier lebt einen Tag. Norma (Männchen, geb. 21.08.1996), Prisca (Weibchen, geb.
14.09.1996) und Dagmar (Männchen, geb. 18.09.1996) ziehen ihr Jungtier
dagegen auf, das am 23.08.1996 geborene Jungtier von Quicka wird von Norma
übernommen
und über 20 Tage (Todestag 13.09.1996) getragen. Besonders hervorzuheben
ist sicherlich, dass nun auch unser altes Wildfangweibchen Gesa seine
Reproduktionsfähigkeit belegte und dass ein Weibchen der Hoechster Totenkopfaffen
erfolgreich züchtete.
Während dieser Zeit nahmen auch vorübergehend wieder Angriffe auf Weibchen der
Tito-Untergruppe zu. Nach einigen Monaten „ohne beobachtete agonistische
Interaktionen unter den beiden Weibchen-Untergruppen tritt nun eine auffällige
Veränderung auf: Eines der Weibchen der ehemaligen (Nur-Weibchen)-Gruppe hat
eine Bißwunde am Kopf, ... Die räumliche Aufteilung der Tiere ist nun ebenfalls
verändert. Die Individuen der (Nur-Weibchen-Untergruppe) nehmen nur noch weniger
als ein Drittel des Geheges ein, während die Weibchen der Tito-Untergruppe sich im
restlichen Raum frei bewegen. ... Es werden immer wieder einzelne Angriffe und
Verfolgungen seitens der vormals bedrängten Tito-Untergruppe auf mehrere Weibchen
der (Nur-Weibchen)-Untergruppe beobachtet, die sich nun meist dichtgedrängt
auf einem Brett aufhalten und vor den Tito-Weibchen weichen. Alle adulten
Weibchen der Tito-Untergruppe (mit Ausnahme von Cindy, die erneut die
Untergruppe gewechselt hat und im Januar 1997 zurück zur Tito-Untergruppe
gewechselt ist) initiieren agonistische Interaktionen. Im Februar 1997 beginnt
eine weitere Paarungsphase. Ende Februar muss Jansen ... separiert werden.
([209], Seiten 162 - 163). Zeitgleich stirbt Oma, das wohl älteste der Hoechster
Weibchen.
Pia Steinweg versuchte nun, die unterlegene Nur-Weibchen-Untergruppe zu
stabilisieren und reintroduzierte die am 20.03.1996 separierten Weibchen Sira und
Hulda. Diesen beiden am 13.03.1997 introduzierten Weibchen gelang es aber nicht,
Anschluss an ihre alte Weibchenuntergruppe zu finden. Pia Steinweg beschreibt
ausführlich diesen am 16.03.1996 beendeten Versuch, teilweise erfolgten die
Angriffe der Tito-Untergruppe so vehement und so schnell, dass ein exaktes
Protokollieren unmöglich war. Ohne einen Alterstod ausschließen zu wollen,
starben beide Weibchen nach diesem Versuch nach meiner Vermutung an
Stress. Am 24.03.1996 versuchte sie eine Entlastung der Situation durch das
Verbringen der im März 1996 separierten Männchen Muck und Yatzee in den
Nachbarkäfig, mit diesem Vorgehen hatte sie einen scheinbar spontanen Erfolg. „Die
Individuen der bedrängten Gruppe machen bald einen entspannteren Eindruck
und verteilen sich etwas großräumiger. Die Weibchen der Tito-Untergruppe
sammeln sich nach einiger Zeit unter zunehmender Vokalisation, wobei die beiden
jungen Männchen anvisiert werden. ... Agonistische Interaktionen innerhalb
der Weibchengruppe werden nur noch vereinzelt beobachtet. ... Etwa drei
Wochen nach der Verbringung der beiden Männchen in das benachbarte Gehege
beginnen jedoch von neuem agonistische Interaktionen. ... Um keine weiteren
Verluste
zu riskieren, werden die beiden Untergruppen am 08.05.1997 getrennt.“ ([209], Seiten
166 - 167).
Vorausgegangen waren diesem Trennungsbeschluss Beobachtungen, die ich - wieder
einmal im Amt - bei den bolivianischen Totenkopfaffen am 06.05.1996 machen
musste/durfte. Die schon ramponierte Prisca saß passiv auf einem der unteren
Ruhebretter. Die beiden kleinen Männchen der Tito-Untergruppe näherten sich ihr
langsam und zupften an ihr, anschließend kehrten sie zu ihrer Untergruppe zurück.
Minuten später näherte sich die gesamte Untergruppe der armen Prisca, die jungen
Männchen - eigentlich viel zu alt zum Getragenwerden - lagen auf dem Rücken ihrer
jeweiligen Mutter und massierten ständig deren Brüste, dann fielen alle erwachsenen
Weibchen über Prisca her. Ich vermutete, den Grund für die Überlegenheit der
Tito-Untergruppe beobachtet zu haben: Die jungen Männchen setzten ihre Mütter ein
wie Kampfroboter. Ich ließ Prisca separieren und empfahl Pia Steinweg, das
Fusionsexperiment abzubrechen.
Die Fusionsexperimente haben deutlich gezeigt, dass sich auch bolivianische
Totenkopfaffenweibchen feindlich gegenüber fremden Weibchen verhalten. Zudem
scheinen sie sich zu merken, welche Tiere die „Bösen“ sind. Diese feindlichen
Beziehungen können in gewissen Grenzen durch gemeinsame Haltung und daraus
resultierender Vertrautheit gemildert werden. Haben sie aber die Chance
zurückzuschlagen, fehlt ihnen die Einsicht, dass ihre Gegner Artgenossen sind. Zur
hohen Bedeutung verwandtschaftlicher Beziehungen berichte ich noch über Pia
Steinwegs letztes Experiment vor dem plötzlichen Verschwinden ihrer Versuchstiere.
Nach dem Abbruch des Experiments wurden 1997 noch (von Jansen gezeugte)
Jungtiere geboren, zwei weibliche Jungtiere von Cindy (05.07.1997) und Yvonne
(07.07.1997) und ein männliches von Leni (13.07.1997), wobei nur das Kind von Leni
auch aufgezogen wurde. Cindy wurde nach der Geburt in der Gruppe verfolgt und am
09.07.1997 separiert.
„Da Cindy nach einer Woche vollkommen regeneriert erscheint, wird sie am
16.07.1997 mit dem Weibchen Prisca fusioniert. ... Nach einem kurzen und wenig
heftigen aggressiven Kontakt, der von Prisca ausgeht, leben die beiden Weibchen über
zehn Wochen friedlich nebeneinander, affiliative Kontakte werden jedoch nicht
beobachtet.
Am 03.09.1997 werden diese beiden Weibchen ... mit dem jungadulten Männchen
Yatzee fusioniert, was ohne agonistische oder affiliative Kontakte verläuft. Prisca
nimmt in den folgenden Wochen wiederholt Kontakt zum Männchen auf. Am
21. bzw. 23.10.1997 werden diese drei Tiere mit zwei adulten Weibchen der
Nur-Weibchen-Gruppe (Norma und Dagmar) fusioniert. Es kommt rasch zu affiliativen
Kontakten (Kontaktsitzen und Fellpflege) zwischen Cindy und ihrer Mutter
Norma und ihrer Schwester Dagmar. Zwischen dieser Matrilinie und der aus der
Nur-Weibchen-Gruppe vertrauten Prisca werden weder agonistische noch affiliative
Interaktionen beobachtet. ... (Am 03.11.1997) wird Priscas 1996 geborene Tochter
Elena in die Gruppe eingeführt. Mutter und Tochter gehen zwar aufeinander zu,
nehmen aber keinen direkten Kontakt auf. Die Anwesenheit des etwa ein
Jahr alten Weibchens ruft zunächst keine deutliche Erregung bei den anderen
Gruppenmitgliedern hervor. Wenige Minuten nach der Fusion nimmt jedoch ... Norma
Kontakt auf und bedrängt das juvenile Tier durch Umklammern. Während Elena
vokalisiert, nähert sich Prisca und hält ihrerseits Norma fest, worauf Elena freigegeben
wird. Eine ähnliche Situation wiederholt sich kurze Zeit später. Daraufhin wird kein
agonistisches Verhalten mehr beobachtet. Elena und Prisca sitzen nun häufig
in affiliativem Kontakt und Elena bleibt im Verband.“ ([209], Seiten 167 -
168).
Ich habe zu Beginn dieses Kapitels berichtet, dass Fremdelternpflege/Tantenverhalten bei unseren
Totenkopfaffen kaum zu beobachten war (s. o.). In den von Pia Steinweg ([209]) und Irmtrud
Winzek
beobachteten Gruppen - Irmtrud Winzek beobachtete die Viktor-Gruppe und
die Karl-Gruppe, Pia Steinweg alle von uns gehaltenen Totenkopfaffen - war
Tantenverhalten häufig zu beobachten. Die Tanten entstammten der Untergruppe der
jeweiligen Mutter. Zwei typische Tanten waren zu unterscheiden, nämlich nullipare
Weibchen und laktierende Weibchen,
die ihr eigenes Jungtier verloren hatten (verloren durch Tod oder durch das
routinemäßiges Entfernen junger Männchen). Letzere ließen die Jungtiere auch saugen.
Die Anzahl der pflegebereiten Tanten war dabei größer als die Zahl der aktiven
Tanten. Potentielle Tanten boten sich den Jungtieren durch eine charakteristische
Körperhaltung an, das jeweilige Jungtier entscheidet dann, ob es die Einladung
annimmt oder nicht. Mütter dagegen fordern ihre Jungen nicht zum Aufsteigen auf, sie
werden von diesen einfach aufgesucht. Insofern kann man nach dem Verhalten Mütter
und Tanten eindeutig unterscheiden.
Nach unseren Untersuchungen darf davon ausgegangen werden, dass
Totenkopfaffen-Gruppen grundsätzlich Weibchengruppen sind, die aus verschiedenen
Untergruppen bestehen, beruhend auf mütterlicher Verwandtschaft. Insofern dürften
auch im Freiland die Weibchen einer solchen Gruppe nahe verwandt sein.
Zwischen diesen Untergruppen treten auch erhebliche Konflikte auf, die in
der Regel aber nicht augenscheinlich sind. Totenkopfaffenweibchen können
auch mit feindlichen Weibchen zusammenleben, doch tradieren sie diese alten
Feindschaften, da positiv soziale Beziehungen in größeren Sozialgruppem auf
Untergruppenmitglieder beschränkt sind. So „vererben“ sie gewissermaßen
Freundschaft und Feindschaft.
Die positiv sozialen Beziehungen werden durch engen Körperkontakt der Beteiligten
offensichtlich. Totenkopfaffenweibchen einer Untergruppe bilden sogenannte
Kuschelgruppen oder „huddling groups“ die von Sigrid Hopf ausführlich diskutiert
werden ([93]). Wir konnten diese engen Kontakte aller Untergruppenmitglieder
täglich beobachten, besonders auffällig bei dem Aussperren der Gruppen zum
Zweck der Reinigung des Innengeheges. Unsere große Viktorgruppe hatte zwei
teilweise getrennte Außengehege zur Verfügung. Gerade bei kälterer Jahreszeit
saßen die zwei Untergruppen wie aufgereiht jeweils Tier neben Tier auf einem
höher gelegenen Ruhebrett in verschiedenen Gehegen. Die Untergruppe der
Männchen hingegen saß - auch bei größerer Kälte - dicht gedrängt auf dem
zweifellos unattraktiven Boden. Besonders bei den bisher als Saimiri sciureus
beschriebenen Varietäten ist die strikte Trennung zwischen Männchen und
Weibchen durchgängig. Männliche Kinder müssen die Untergruppen der Mütter
verlassen, sie finden dann Anschluss an andere Männchen, bzw. verlassen
(unter Freilandbedingungen) die Geburtsgruppe. Die Mehrheit der Männchen
dürfte dieses Verlassen nicht überleben, doch werden einige Anschluss an
fremde Männchenuntergruppen finden. In der Reproduktionszeit sind diese
fremden Männchen dann besonders attraktiv als Sexualpartner, finden Zugang
zu den Weibchenuntergruppen und werden anschließend von den Weibchen
verjagt.
Unter Freilandbedingungen finden wir häufig eine Geburtensaison. Dafür
werden in der Literatur verschiedene Gründe angegeben. Da die Freilandbiotope
zu unterschiedlicher Jahreszeit mehr oder weniger Nahrung bieten, wird als
der die Saisonalität bedingende Faktor das Nahrungsangebot angenommen.
Je nach den Bedingungen der untersuchten Art, also den unterschiedlichen
Lebensräumen, wird dann spekuliert, dass entweder die Jungen nur bei hohem
Nahrungsangebot die Aufzuchtsphase überleben oder dass die Männchen nur bei
hohem Nahrungsangebot die Strapazen der Reproduktionszeit durchstehen
können. Unter den Bedingungen in Menschenobhut gehen diese Saisonalitäten
verloren. Dies gilt aber nicht für Totenkopfaffen. Hier bleibt das saisonale
Reproduzieren auch in Menschenobhut erhalten. Oestrische Weibchen rufen
offensichtlich bei anderen Weibchen einen Oestrus hervor. Insofern ist nicht
verwunderlich, dass paarweise oder in Kleinstgruppen gehaltene Totenkopfaffen
nicht züchten. Die saisonale Fortpflanzung ist also ein angeborenes Merkmal
der Weibchen. Unter welchen ökologischen Bedingungen bzw. unter welchem
Predationsdruck dieses Verhalten in den Millionen Jahren der Entwicklung
selektioniert wurde, ist reine Spekulation, an der ich mich nicht beteiligen
möchte.
Die oben aufgezeigten Verhaltensunterschiede zwischen bolivianischen Totenkopfaffen
und Saimiri sciureus konnten auf breiter Basis belegt werden. „Während (die soziale
Fellpflege) bei S. sciureus nur selten zu beobachten ist, kommt sie bei S. boliviensis
relativ häufig vor.“ ([209], Seite 155). Steinweg stellte auch die Kontakte
zwischen Männchen und Weibchen als diskriminierende Komponente vor.
Doch sind diese sozialen Kontakte zwischen den Geschlechtern keineswegs ein
zwingender Vorteil, weit eher eine Lebensgefahr für die bolivianischen Männchen,
ändern diese doch nichts an dem Verhalten der Weibchen, vertraute Männchen
grundsätzlich zu verjagen, um nach fremden Ausschau zu halten. Zudem
belegte Steinweg Unterschiede zwischen den beiden Formen bei dem Genitalen
Imponieren.
Nach der neuesten Literatur ([141]) werden auch bei den Totenkopfaffen zahlreiche Arten
beschrieben. Unsere Tiere aus British Guayana wären weiterhin Saimiri sciureus, die
bolivianischen Totenkopfaffen Saimri boliviensis boliviensis, die Tiere der Viktorgruppe
Saimiri cassiquiarensis und die von Baldwin in Florida untersuchten Tiere Saimiri
boliviensis peruviensis. Es mag für diese Unterteilung gute Gründe geben, doch sind
nach dem Verhalten die Totenkopfaffen aus British Guayana, Kolumbien und Peru eine
mehr oder weniger einheitliche Gruppe, denen die bolivianischen Totenkopfaffen
gegenüber stehen. Damit komme ich im Prinzip zu der gleichen Auffassung wie
Thorington 1985 ([225]) bei seiner sorgfältigen Analyse der Daten zum System der
Totenkopfaffen.
Im Gegensatz zu ihm würde ich aber statt von einer einzigen Unterart von
einer einzigen Art sprechen. Soweit es gute Gründe geben sollte, diese Art
auf Artenniveau aufzuspalten, dann erscheint es mir zwingend, auch Saimiri
boliviensis boliviensis und Saimiri boliviensis peruviensis auf Artenniveau zu
trennen.

Impressum/Kontakt