Abbildung 13.1: Die Pionierin der Kapuzineraffenforschung Angela Nolte bei
einem gemeinsamen Besuch im Nürnberger Zoo
Die bisher in diesem Buch besprochenen Affenarten waren nicht meine „Wunschaffen“,
vielmehr habe ich mit und an diesen nur gearbeitet, weil sie in der von mir betreuten
Primatenstation vorhanden waren. Ganz anders war die Wahl der Kapuzineraffen als
Versuchsobjekte.
Kapuziner1
waren schon früh als Leierkastenaffen und als Versuchstiere für verschiedenste
Lernexperimente den Europäern vertraut. Um so mehr erstaunt es, dass in den letzten
Jahrzehnten an Kapuzinern so gut wie überhaupt nicht gearbeitet wurde, galten
doch gerade Vertreter dieser Species als besonders lernwillig und intelligent.
„Modeaffen“ der Wissenschaft waren vielmehr der Rhesusaffe Macaca mulatta und in
den letzten Jahrzehnten der Totenkopfaffe Saimiri sciureus. Mein Doktorvater
Heinrich Sprankel berichtete in seinen Vorlesungen zur Primatologie über die
erstaunlichen Leistungen der Kapuzineraffen und stellte uns Ergebnisse vor, die
Angela Nolte 1958 ([149]) publiziert hatte (über das Instinktverhalten der
Kapuzineraffen). Er schloss seinen Bericht mit der Aussage: „Irgendeiner
müsste einmal Kapuzineraffen genauer untersuchen.“ Diese Aufforderung nahm
ich an und etablierte in Kassel eine Kolonie des Gehaubten Kapuzineraffen
Cebus apella. Über die Schwierigkeiten und Erfolge werde ich in diesem Kapitel
berichten.
Kapuzineraffen sind zu ihrem Namen gekommen, da Carl von Linné bei der
Erstbeschreibung eines Weißschulteraffens Cebus capucinus Ähnlichkeiten mit einem
Kapuzinermönch erkannt haben wollte (nach der Haartracht). Unsere Kapuzineraffen
haben zusätzlich eine auffällige Haartracht, die man als Haube ansprechen mag.
Während der Weißschulteraffe seinen Namen behalten hat, haben unsere
Kapuzineraffen ihren wissenschaftlichen Namen verloren. Die größeren Kapuzineraffen
werden in der neuesten Systematik ([141]) einer eigenen Gattung Sapajus zugeordnet
mit einer Vielzahl verschiedener Arten, so dass meine Kapuzineraffen heute
nicht mehr Cebus apella sind. In diesem Beitrag behalte ich aber den alten
Namen bei, da ich mit diesem zahlreiche Arbeiten veröffentlicht habe und durch
einen neuen Namen meine eigenen Beiträge „abwerten“ würde. Ich gebe aber
die jeweils heute gültigen Namen an, so dass meine Leser sich orientieren
können.
Bevor ich mit meinem Bericht beginne, möchte ich einige der von Angela Nolte
erkannten angeborenen Verhaltensweisen der Kapuzineraffen vorstellen. Zum
Aussaugen von Früchten führte sie aus: „Der Kopf wird dabei weit nach hinten gelegt,
und die Früchte werden zwischen Zunge und Gaumen, der mit starken Querleisten
versehen ist, ausgedrückt. Die Schalen, Fruchtfleischreste oder Kerne spucken die Affen
aus“ ([149], Seite 185). Erstmals beschrieb Nolte das Tischchenmachen: „Diese
Verhaltensweise tritt besonders regelmässig beim Fressen von Körnern auf. Die
Affen nehmen gern den Mund voll Körner, spucken den Inhalt auf die eng
nebeneinander gelegten Unterarme und fressen die Körner nacheinander ab. Mit
dieser Tischchen-Reaktion fangen sie auch die bei ihrem recht hastigen Fressen
herunterfallenden Futterbrocken auf. ... Das Tischchenmachen ist meines
Erachtens nach eine angeborene Verhaltensweise der Kapuziner, die in der freien
Natur von grosser Bedeutung sein kann, da die Affen sich vorwiegend in den
Kronen hoher Bäume aufhalten und nur selten am Boden fressend beobachtet
wurden“ ([149], Seite 185). Auch das Einreiben mit Zwiebeln wird von ihr
besprochen: „Gibt man einem Kapuziner eine Zwiebel, so beisst er hinein, zwinkert
mit den Augen, legt sich häufig auf eine Seite und reibt mit den Händen, in
denen er ein Stück Zwiebel hält (oder die nach Zwiebeln riechen), über den
Kopf, über den hinteren Rückenabschnitt bis zur Schwanzwurzel, mit den
Füssen über das Fell des Nackens und der dorsalen Seite der Oberarme. Die
Reihenfolge der Arm- und Beinbewegungen ist variabel. ... Man hat durchaus den
Eindruck, als bereite den Affen dieses Einreiben grosses Behagen“ ([149],
Seite 193). Bei dem Hämmern (der Hämmerer des Waldes) war Nolte sich
noch nicht sicher bei der Bewertung: „Ob hier eine angeborene Reaktion, ein
Lernen durch Erfahrung oder ein einsichtiges Handeln vorliegt, ist bisher nicht
eindeutig entschieden worden“ ([149], Seite 190). Nolte diskutiert auch den
Werkzeuggebrauch. Zu beiden Fähigkeiten der Kapuzineraffen, das Aufhämmern
von Nahrung und den Werkzeuggebrauch, werde ich noch eigene Ergebnisse
berichten.2
Ich konnte diese angeborenen Verhaltensweisen der Kapuzineraffen täglich in meiner
Kolonie beobachten, habe diese aber nicht hinreichend dokumentieren können.
Dies habe ich nun an Enkeln und Urenkeln meiner Kapuzineraffen im Zoo
Augsburg3
für diesen Beitrag nachgeliefert.
Freilich sind weder die Beobachtungen in Kassel noch in Augsburg Beobachtungen im
natürlichen Lebensraum. Diese sind nur schwer zu erbringen. In erreichbaren Gebieten
(Vorhandensein einer Straße) sind Kapuzineraffen ausgerottet, in Südamerka gibt es
keinen Jagdherren, der die Jagd als sein Privileg betrachtet, insofern sind Kapuzineraffen
billige Nahrungsmittel. Jungtiere werden von den Kindern als Spielzeug gehalten, bis
sie zubeißen, dann werden sie an einen Spieß gebunden und geröstet. Insofern
muss man sich Untersuchungsgebiete außerhalb menschlicher Ansiedlungen
suchen4,
manchmal kann man Affen auch auf Farmgelände antreffen, deren Besitzer sich aus
irgendeinem Grund ein Stück Urwald erhalten wollen. Aber auch hier ist
das Beobachten äußert schwierig. Insofern sind meine Befunde solche unter
Luxusbedingungen in Menschenobhut.
In Kassel boten wir unseren Kapuzineraffen all das Leckere, was ein Großmarkt
liefert, an. Sie waren begeistert. Nur zweimal entsetzten wir unsere Tiere. Einmal
schenkten wir ihnen einen großen Weihnachtsmann. Die Affen, die selbst mit einer
wilden Ratte fertig wurden, fürchteten sich und schraken zurück. Schließlich
traute sich Toko, über den ich noch mehrfach berichten werde, beherzt, den
Weihnachtsmann zu greifen und - wie eine Kokosnuss - aufzuschlagen. Die Reste
vertilgte die gesamte Gruppe. Ein anderes Mal hatte meine Mitarbeiterin Yon
Hille uns anlässlich ihres Geburtstages eingeladen und einen leckeren Salat
mitgebracht. Bestandteile dieses Salates waren kleine Babykraken (achtarmige
Tintenschnecken), die aber keiner von uns essen wollte. Wir gaben diese daher unseren
Kapuzineraffen und riefen bei diesen Ensetzen hervor. Nur sehr vorsichtig und mit
„spitzen“ Fingern wagten sie sich an das ihnen unbekannte Futter heran. Diese
unbeabsichtigten Versuche wollte ich gerne in Augsburg wiederholen. Ich
konnte Herrn Reviertierpfleger Peter Kühlburg überzeugen, dass seine Affen
hierdurch keinen Schaden nehmen würden, und er erklärte seine Bereitschaft
mitzuwirken.
Durch diese Unterstützung und durch die Mitwirkungsbereitschaft der Augsburger
Kapuzineraffen, die mich in ihrem Gehege duldeten, konnte ich nun das in Kassel
Versäumte nachholen und hier dokumentieren.
In der Tabelle 13.1 habe ich alle nichtmenschlichen Mitwirkenden
angegeben.5








Der Weihnachtsmann (Abb. 13.18) löste auch in Augsburg Entsetzen (Abb. 13.19) aus.
Das dominierende Männchen Purzel traute sich aber immer wieder an den Tisch
heran, zog und wackelte an diesem (Abb. 13.20), bis der Weihnachtsmann
umfiel. Ein weiteres Wackeln führte zu keinem weiteren Erfolg. Es gab keinen
Toko in der Gruppe (wie in Kassel, s. u.), wir brachen den Versuch ab. Am
nächsten Tag boten wir wieder den nun liegenden Weihnachtsmann, garniert mit
Walnüssen, an. Letztere wurden zwar vorsichtigst Stück für Stück aus dem
„Gefahrenbereich“ entfernt (Abb. 13.21), der Weihnachstmann blieb unberührt. Erst
nachdem Manuela Dietz unseren Weihnachtsmann zerteilte, war die Gefahr gebannt.
An dem Verzehr der Schokolade beteiligte sich dann die gesamte Gruppe (Abb.
13.22).
Bei dem Tintenschneckenversuch verzierten wir das angebotene Futter mit
einer Krake. Diese wurde nicht beachtet und bei der hastigen Futteraufnahme
heruntergeschleudert. Bei dem Angebot einer Futterschüssel nur mit Kraken hatten die
Kapuzineraffen zwar keine Probleme, sich der Schüssel zu nähern (Abb. 13.23), doch
trauten sie sich nicht, eine Krake zu probieren. Wie die Kasseler Kapuzineraffen waren
auch die Augsburger Nachkommen entsetzt. Am nächsten Tag boten wir
die nun sorgfältig abgewaschenen Tintenschnecken erneut an. Nur einer der
Kapuzineraffen, Langer Hans, versuchte das Futter (Abb. 13.26). Erst als
Manuela Dietz sich zu ihnen setzte und selber Krake für Krake aß (Abb.
13.27), war der Bann gebrochen, aus ihrer Hand und ihrem Mund griffen die
Kapuzineraffen nach den Kraken und „genossen“ diese dann auch ohne menschliche
Unterstützung.
Die Kapuzineraffen in Augsburg lernten also, dass Kraken essbar sind. Sicherlich
werden unter Freilandbedingungen ähnliche Prozesse ablaufen. Die Vorsicht vor
Unbekanntem ist zweifellos evolutionär stabil und wird nicht selektioniert.
Durch das mutige Ausprobieren noch nicht bekannter Ressourcen durch ein
Individuum können Futtertraditionen entstehen, die möglicherweise dann
von Gruppe zu Gruppe unterschiedlich sind. Das angeborene Verhalten im
Zusammenhang mit der Nahrungsaufnahme wird dann also an neuen Objekten
praktiziert.
So können Kapuzineraffen sich auch problemlos einen menschlichen Haushalt
erschließen, wodurch sie aber direkte und erfolgreiche Konkurrenten um knappe
Ressourcen werden. Ich bin sicher, unsere Kapuzineraffen hätten sich auch frei in
Kassel behauptet. Sie wären zwar eine Behinderung des Autoverkehrs geworden,
hätten aber sicherlich den nächsten Lebensmittelmarkt gefunden und erobert. Da
Affen bei uns nicht „heilig“ sind, hätten sie dadurch freilich auch das Ende der
„Freihaltung“ bewirkt und wären entweder erschossen worden oder in einer
restriktiven Haltung gelandet.



In Kassel wurde ich von Besuchern regelmäßig gefragt, ob man nicht das Gehege
begrünen könnte. Diesen demonstrierten wir die Antwort. Ich ließ einen Baum holen
und in den Käfig stellen. Nicht umsonst werden die Kapuzineraffen „Gärtner des
Waldes“ genannt, innerhalb weniger Minuten zerschroteten sie diesen Baum. Wir
gaben ihnen auch regelmäßig nicht verkaufte Weihnachtsbäume zum Zerlegen.
Entsprechende Erfahrungen hatte auch Herr Kühlburg gesammelt. Wir boten
also auch den Augsburger Kapuzineraffen einen Baum an. Doch waren die
Augsburger Kapuzineraffen wohl durch mein Vorhandensein oder durch die bereits
durchgeführten Versuche nicht mehr so motiviert. Sie zerstörten ihn erst in den
Folgetagen. Doch demonstrierten sie wunderbar die Leistung des Greifschwanzes im
dünnen Geäst (Abb. 13.29). Gleichzeitig demonstrierten sie, dass das Blattwerk
selbst aus kurzer Entfernung das individuelle Ansprechen erheblich erschwert.
Man kann ermessen, wie schwierig es ist, ein Individuum in 50 m Höhe sicher
anzusprechen.
Zum Abschluss meines Besuches in Augsburg demonstrierte Peter Kühlburg noch zwei
seiner Ideen, das Zusammenleben der Kapuzineraffen attraktiv zu gestalten. Er
sammelt verschiedene Zapfen und bietet diese seinen Kapuzineraffen an (Abb. 13.34).
Diese stießen auf großes Interesse, wurden untersucht und ausgeklopft bzw. auch
kunstvoll zerlegt (Abb. 13.32). Seine Mühen des Sammelns werden durch die
Kapuzineraffen belohnt.
Darüber hinaus bietet er seinen Tieren alte tierreiche Wurzeln an, an denen sich seine
Kapuzineraffen abarbeiten können, sie sammeln Insekten bzw. stochern nach Maden u.
ä. (Abb. 13.35, 13.36)
Zu den Augsburger Namen sei bemerkt, bei der individuellen Unterscheidung einer
größeren Gruppe (noch) nicht bekannter Tiere erleichtert es das Kennenlernen, wenn
man nach irgendwelchen Merkmalen Namen wählt oder diese nach ihnen (angeblich)
ähnlichen Menschen benennt. Frau Matys (Kapitel 4) sah eben aus wie Frau
Matys, auch wenn sie nur eine Galago-Dame war. Der kleine Christian verriet
während meiner Aktivitäten sein Geschlecht. Man fragte mich, ob ich Einwände
hätte, ihn nach mir zu benennen. Dieses Ansinnen habe ich als Auszeichnung
empfunden und gerne mein Einverständnis erteilt. In der Regel habe ich das
Namengeben meinen Mitarbeitern überlassen, bzw. auch nach ihnen die Affen
benannt.
Da Peter Kühlburg und Manuela Dietz die Individuen auf meinen Aufnahmen
identifizierten - nur bei den Baumaufnahmen mussten sie passen -, wird augenfällig,
dass neben Purzel und Clown Kasper auch Langer Hans ein besonders kooperierender
Kapuzineraffe ist.



Bei unserer Arbeit an und mit Kapuzineraffen war ich an den Befunden zum sozialen
Zusammenleben besonders interessiert. Andere Fragestellungen, wie z. B. zum
Kommunikationsverhalten, wären sicherlich auch von großem Interesse, doch unter
unseren Bedingungen - Leben in einem großen Sozialverband - waren diese
Untersuchungen nicht zu leisten. Einmal wollte ich z. B. die Lautäußerungen unserer
Weibchen im Oestrus mit einem extra hierfür angeschafften Richtmikrofon aufnehmen.
Diese extrem auffällige und auch von noch nicht trainierten Beobachtern leicht zu
hörende und eindeutig zuzuordnende Vokalisationsform war nicht aufnehmbar, sie
wurde überlagert durch den Haltungslärm. Kapuzineraffen geben ständig
Stimmfühlungslaute von sich und machen „Krach“ bei der schnellen Fortbewegung im
Gehege, solche Geräusche blendet der menschliche Beobachter mit seinem Gehör
einfach aus, die Tonaufnahme dagegen vermittelt nur ungeheuren Lärm. Selbst das
„Begrüßungsgeschrei“ (s. u.) konnte ich nicht aufnehmen, begrüßte mich doch selbst
Bubi nicht (s. u), wenn ich mich mit dem Mikrofon in der Hand dem Käfig
näherte.
Anderen Kommunikationsformen fehlte die Eindeutigkeit. So könnte ich auch
über das Genitale Imponieren (vgl. Kapitel 11) bei Kapuzineraffen berichten,
doch habe ich diese Verhaltensweise nicht als Imponieren empfunden.
Bei hoher Erregung erigieren Männchen den Penis und Weibchen die
Klitoris6,
dies war mein banaler Befund. Insofern haben wir das Genitale Imponieren und auch
das auffällige Harnwaschen (Abb. 13.37) überhaupt nicht protokolliert. Darüber hinaus
fehlte häufig die Eindeutigkeit der Verhaltensweisen. So konnten wir z. B. bei dem
Drohen (Abb. 13.38), das schwache Drohen, das starke Drohen und das starke Drohen
mit Lautäußerung leicht unterscheiden. Unsere Kapuzineraffen differenzierten
aber anders, sie entschieden in Abhängigkeit von der sozialen Position des
Sozialpartners. Das schwache Drohen - ein für einen nicht geübten Beobachter kaum zu
erkennendes leichtes Zurückziehen der Mundwinkel - eines dominierendes Weibchens
z. B. führt zur panischen Flucht des betroffenen Individuums. Das starke
Drohen mit Lautäußerung, eindrucksvoll und imponierend, eines großen, aber
„unbedeutenden“ adulten Männchens hingegen wird von allen Sozialpartnern
ignoriert.
Nach Anfangsschwierigkeiten, über die ich noch berichten werde (s. u.), war
unsere Kapuzinerhaltung sehr erfolgreich. Ich wertete gemeinsam mit meiner
Mitarbeiterin Annette Klaiber-Schuh ([264]) 1995 auch bei den Kapuzineraffen die
Reproduktionsdaten unserer Kolonie aus. Insgesamt wurden bis zum Zeitpunkt der
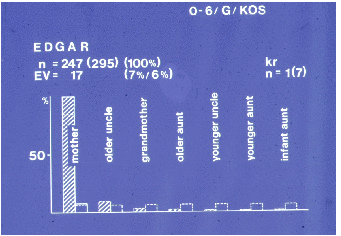
Auswertung 188 Kinder (in der Regel Einlinge, nur eine Zwillingsgeburt) geboren. 62
% der Neugeborenen überlebten die erste Lebenswoche, von den Überlebenden
erreichten dann 85 % zumindest das erste Lebensjahr. Erstgebärende Weibchen (n =
22) waren weniger erfolgreich als mehrfachgebärende, nur 24 % der Kinder
überlebten die erste Lebenswoche. Die zweitgeborenen Jungtiere (n = 18) hatten
schon höhere Überlebensraten (44 %), erst aber der dritten Geburt hatten die
Neugeborenen die gleichen Chancen wie die Kinder der erfahrenen Wildfangweibchen.
Das mittlere Alter der erstgebärenden Weibchen war 5 Jahre 8 Monate (± 3
Monate). Der Geburtenabstand hängt deutlich von dem Aufzuchtserfolg ab, er
betrug 603 ± 21 Tage bei Kinder tragenden Weibchen (n = 88) und 338 ±
14 Tage bei Weibchen, die das Kind verloren hatten (n = 72). Der kürzeste
Abstand zwischen zwei Geburten betrug 157 Tage. Eine Saisonalität war bei den
Geburten in der Kolonie nicht auffällig, die meisten Kinder (n = 156) wurden
zwischen April und September geboren, unabhängig von der Haltungsdauer
in unserer Kolonie und unabhängig von der Vorgeschichte (Wildfang oder
Nachzucht).
Das Geschlechterverhältnis Männchen zu Weibchen betrug 1 : 0,90 bei allen
Jungtieren, 1 : 0,80 bei den überlebenden Kindern. Die höhere Überlebensrate
männlicher Kinder bzw. die höhere Mortalitätsrate weiblicher Kinder war
auffällig.
Die Hauptträgerin der Kinder ist die eigene Mutter. Soweit Fremdelternpflege
überhaupt beobachtet werden konnte (ab der vierten Lebenswoche), war das Tragtier
in der Regel eine Verwandte/ein Verwandter der Mutter oder deren „Freund“ (s.
u.).
Den Bericht über unsere Kapuzineraffenergebnisse unterteile ich in zwei Teile.
In einem ersten Teil berichte ich über die Ergebnisse in der provisorischen
Primatenhaltung (1975 bis 1984) und in dem zweiten Teil über die Erkenntnisse, die
wir ab 1985 in der fertig gewordenenen Primatenstation gewinnen konnten.
An der Erkenntnisgewinnung waren zahlreiche Studentinnen und Studenten
engagiert beteiligt. Drei Dissertationsarbeiten ([153], [115], [39]) trugen dazu bei,
zahlreiche Forschungsfragen zu beantworten. Einige Ergebnisse der Arbeiten von
Petra Pippert und Hella Kröger geb. Höhmann, die auch als wissenschaftliche
Mitarbeiterinnen maßgeblich die Beobachter trainierten und das Beobachten
organisierten, werden Bestandteil dieses Kapitels sein. Die Staatsexamensarbeiten
liegen mir leider nur teilweise vor, die Themen derjenigen von Heike
Roland7 und
Gabriele Werner8
habe ich aus Pippert ([153]) ungeprüft übernommen.
Die Staatsexamensarbeiten von Claudia
Brinkmann9,
Petra Pippert10,
Sabine Schulz11,
Michael Dulitz12,
Katharina Klewitz13,
waren jeweils wichtige „Meilensteine“. An den Befunderhebungen waren auch
Karin Bourgeois, Beate Holstein, Dagmar Lentzkow, Uwe Orth und Cornelia
Schäfer-Witt beteiligt, ihnen und den möglicherweise nichterwähnten Mitarbeitern sei
ausdrücklich für ihr Engagement gedankt, das das Projekt „Kapuzineraffen“ zu
einem erfolgreichen werden ließ. Dank der Förderung durch die Deutsche
Forschungsgemeinschaft hatte ich auch genügend Personal- und Sachmittel, um die
Kapuzineraffen täglich beobachten lassen zu können. Wir protokollierten täglich
das Verhalten der Sozialgruppen und zusätzlich zahlreiche Individuen über
„Einzeltierbeobachtungen“- nur ein Individuum stand im Fokus des Beobachters -, zudem
filmten14
wir regelmäßig das Zusammenleben der Kapuzineraffen. Insofern wurden
die Kapuzineraffen seit dem Beginn der Förderung durch die Deutsche
Forschungsgemeinschaft täglich zumindest drei Stunden lang beobachtet. Von dem
vorliegenden Datenmaterial stelle ich hier exemplarisch einige Ergebnisse vor. In der
Zeit, in der ich die Primatenstation geleitet habe, konnte ich nur einen Teil der
vorliegenden Daten auswerten lassen. Weitere detailliertere Analysen unser
Beobachtungen werde ich vielleicht in der Zukunft vorstellen können. Hierfür hat
mir die „Adelheid-Welker-Stiftung zur Förderung der primatenethologischen
Forschung“ Personalmittel in Aussicht gestellt.
Bei meinem Wechsel nach Kassel 1974 waren eigentlich keine geeigneten
Haltungsbedingungen vorhanden. Der Leiter der Arbeitsgruppe Zoologie
und vergleichende Anatomie, Prof. Dr. Werner Meinel, hatte für seine
Pläne, eine Primatenhaltung für anatomische Versuchsvorhaben zu
etablieren, vier Räume über den Zentralen Werkstätten der Hochschule
organisiert.15
Diese eigentlich typischen Büroräume mit abgehängten Klimadecken waren zumindest
gefliest und hatten einen Bodenabfluss. In einem Raum, dem späteren Kapuzinerraum,
standen auch bereits drei Käfige vor der Fertigstellung. Bei unseren Gesprächen ging
er auch bereitwillig auf mein Anliegen ein, Kapuzineraffen anzuschaffen. Er bestellte
umgehend „kleine“ Kapuzineraffen bei einem Importeur, er dachte dabei aber wohl an
kleinere Arten.
Bereits am 13. Mai 1974 war es dann soweit, die ersten Kapuzineraffen kamen in
Frankfurt an. Doch unsere Freude war mehr als getrübt, auf dem Transport bzw. in
den ersten Tagen nach Ankunft starben von den 12 importierten Tieren 10, Tschiggo
und Purzel überlebten, wenngleich zunächst in jämmerlichem Zustand. Die Exporteure
hatten uns - so wissen wir heute - Kapuzinerbabies geschickt, die wegen ihres Alters an
sich schon kaum Überlebenschancen gehabt haben. Zudem kamen die Tiere
gemeinsam verpackt in einer kleinen Kiste (um Transportkosten zu sparen), durch
Futterreste, Urin und Exkremente völlig verklebt, an, so dass bereits bei der
Ankunft in Frankfurt der Großteil der Affen tot oder irreparabel geschädigt war.
Die Entscheidung, wie im Tierhandel üblich, die „Sendung“ zurückgehen zu
lassen, konnte ich nicht treffen, hätte ich doch auch die Überlebenden dem
sicheren Tod ausgeliefert. Auch der zweite (Ersatz-) Transport (22.6.1974)
bestand wiederum mehrheitlich aus Babies, von denen ebenfalls nur zwei
überlebten.16
Tschiggo und Purzel hielten wir in demselben Raum, in dem bereits ein von mir aus
Gießen mitgebrachtes Riesengalagopaar (Werner und Ilse) lebte. Ich habe schon
berichtet (Kapitel 4), dass deren Tochter Josephine ihren Käfig mühelos verlassen
konnte, sie „besuchte“ auch regelmäßig Tschiggo und Purzel in deren Käfig. Bei
Ankunft des zweiten Transportes waren bereits Käfige in dem späteren Kapuzinerraum
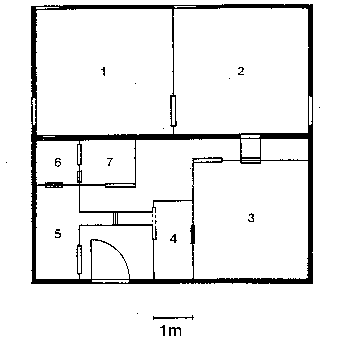
installiert (Käfig 3 und 4 der Haltungsskizze, der Käfig 3 war durch eine Eternitwand
unterteilt). Im hinteren der beiden Teilkäfige lebten ab dem 21.06.1974 drei
Javanermakaken, die sich Werner Meinel von den Behringwerken in Marburg
schenken ließ. In Kapitel 14 werde ich über unsere Haltungserfahrungen und
über Schwierigkeiten berichten. Ich konnte glücklicherweise Werner Meinel
überzeugen, dass die von ihm geplanten Käfige ungeeignet waren. Es fehlten jegliche
Absperrmöglichkeiten, zudem war die gewählte Maschenweite für die geplante
Affenhaltung zu weit. Die Mehrheit der anzuschaffenden Arten hätte die Käfige
mühelos durch das Gitter verlassen können. Er unterstützte auch mein Bemühen,
zusätzlich Außengehege (auf dem Dach des Werkstattgebäudes) errichten zu lassen.
Die Tiere des zweiten Importes konnten in den Käfig 4 (vgl. Abb. 13.44) überführt
werden. Wir hielten also die beiden Paare Kapuzineraffen (Tschiggo/Purzel
und Hoppediz/Eda) räumlich getrennt in verschiedenen Bereichen der
Hochschule.17
Nach der Fertigstellung der Umbauten im Kapuzinerraum (zwei Außenkäfige, drei zusätzliche
Innenkäfige (Käfig 518
und 6 der Haltungsskizze) und nach der Errichtung der Käfige im Makakenraum (mit
einem engeren Käfiggitter - Josephine hatte Meinel überzeugt) besuchten meine Frau
und ich meinen Freund Dr. Iggo Tholen, der damals im Kronberger Opelzoo arbeitete.
Bei einem Rundgang berichteten uns die Tierpfleger, denen ich von meinen
Kapuzinerplänen berichtete, sie hätten einen siebzehnjährigen Kapuzinermann in
Pflege. Die Besitzerin sei wohl verstorben, wenn ich wollte, könnte ich ihn mitnehmen.
So traf ich Bubi. Bubi lebte in einem kleinen „Affenkäfig“, den wohl Privatleute
damals für hinreichend hielten. Die Vorbesitzerin hatte ihm die Eckzähne ziehen
lassen. Durch die jahrelange Haltung unter diesen Bedingungen, vermutlich auch durch
Vitaminmangel, konnte Bubi kaum springen, seine Hüftgelenke waren versteift, Bubi
war viel größer als unsere Kapuzineraffen, seine Fellfarbe jedoch identisch.
Eigentlich war Bubi ein Affe, den ich von seinem körperlichen Zustand her nicht
„gebrauchen“ konnte. Er tat mir aber Leid und offensichtlich hegte er auch Sympathie
für mich. Eine jahrelange Freundschaft begann. Dass gerade Bubi uns so
viele Ergebnisse liefern sollte (s. u.), ahnten wir zu diesem Zeitpunkt noch
nicht.
Wir nahmen ihn am 20.03.1975 mit nach Kassel, ich ließ ihn einen gerade
fertiggewordenen Käfig des noch nicht genutzten Makakenraumes beziehen. Meiner
Tierpflegerin Edith Lorenz hinterließ ich die schriftliche Nachricht, ich hätte einen
zahmen Kapuzineraffen, namens Bubi, mitgebracht. Sie solle am nächsten Morgen auf
mich warten. Hier folgte Edith Lorenz mir nicht, vielmehr rief sie mich am nächsten
Morgen an: „Ihr Bubi hat mich ins Bein gebissen, Scheiße!“. Bubi war frei. Kaum
betrat ich den Raum, begrüßte er mich mit lautem Geschrei und ließ sich mühelos
(mit Unterstützung eines Schrubbers) zurück in seinen Käfig drängen. Durch unsere
vitaminreiche Diät und den größeren Bewegungsraum wurde er auch von
Tag zu Tag gewandter und bewegungsfreudiger und nutzte alle Bereiche des
Käfigs.
Nun war es mein Anliegen, unsere getrennt gehaltenen Kapuzineraffen
zusammenzuführen. Die sozialen Beziehungen bei beiden getrennt beobachteten Paaren
(Tschiggo und Purzel, Hoppediz und Eda) beschränkten sich auf gemeinsames Ruhen
in engem Körperkontakt. Gegenseitige soziale Körperpflege konnte nicht beobachtet
werden, agonistische Interaktionen waren selten. Bei erstem optischen Kontakt aller
vier Tiere war bemerkenswert, dass die mit Hilfe einer Umsetzkiste in einen
benachbarten Käfig überführten Tschiggo und Purzel in ihrer Kiste wie tot verharrten,
während Hoppediz und Eda explorierten. Als jedoch nach 32 Minuten Tschiggo und
Purzel ihre Umsetzkiste vorsichtig verließen, suchten Hoppediz und Eda fluchtartig
einen höhergelegenen Ruheplatz auf und nahmen dort Demutstellung ein.
Am 03. Juni 1975 vergesellschafteten wir die beiden Paare. Alle vier Tiere
suchten zwar bei Ruhe bzw. Gefahr den gegenseitigen Körperkontakt, doch
blieb das völlige Fehlen sozialer Körperpflege auffällig. Nach Separierung des
schwächlichen Weibchens Eda ließen wir schließlich am 06. Juli 1975 auch Bubi hinzu.
Sowohl Bubi als auch das Trio Tschiggo, Purzel und Hoppediz drohten sich
gegenseitig und flohen voreinander. Doch bereits am Abend des Fusionstages
beobachteten wir, dass Bubi den seines Weibchens „beraubten“ Hoppediz putzte,
wobei dieser das Putzen anfänglich nur kurz zuließ und dann floh. Bereits am
folgenden Tag zeigte Bubi soziale Körperpflege auch gegenüber Purzel und
Tschiggo. In den nächsten Tagen waren auch von Tschiggo, Purzel und Hoppediz
ausgehende Anfänge der sozialen Körperpflege, nämlich vorsichtiges Zupfen
an den Haaren der anderen Tiere, zu beobachten; Bubi nahm häufig eine
Putzaufforderungsstellung ein, doch blieb seine Aufforderung meist ohne Antwort. In
der so gebildeten Ausgangsgruppe (A-Gruppe) waren agonistische Aktivitäten selten.
Die Beobachtungen riefen bei uns damals den Eindruck hervor, soziale Körperpflege
müsse gelernt werden. Diese Hypothese konnten wir später eindeutig falsifizieren
(s. u.), unsere Kapuzineraffen waren zu jung, um soziale Körperpflege zu
zeigen.
Die schwere Entscheidung, ob es überhaupt zu verantworten sei, noch weitere
Kapuzineraffen zu importieren, hatte ich bereits vor der Übernahme von Bubi
getroffen. Voraussetzung war die Verbesserung der Importbedingungen. Gemeinsam
mit den Kasseler Veterinären, von denen ich hier Herrn Dr. Primus besonders
erwähnen möchte, habe ich viele Eingaben gemacht, damit sich die Situation bei dem
Import bessert. In den Einfuhrbestimmungen steht nämlich eindeutig, dass nur zwei
Tiere pro Abteil gesendet werden dürfen. Diese Bestimmung einzuhalten und zu
überprüfen, haben wir eindringlichst angeregt. So unerfreulich der Tod der jungen
Affen war, so erfolgreich war dann unser Bemühen, wurden doch dann in
der Folgezeit die Bestimmungen eingehalten, was unzähligen importierten
Affen (nicht nur in Kassel) das Leben gerettet hat. Dafür sei den Herren
vom Staatlichen Veterinäramt in Kassel bzw. den Herren Veterinären beim
Regierungspräsidenten in Kassel besonders gedankt. Wir forderten nicht nur
nachdrücklich, dass die Importbestimmungen eingehalten werden und die Tiere
adult19
sein sollten, wir installierten auch in mühevoller Kleinarbeit für alle Fälle verstellbare
Zwischenböden und -decken (Fichtenholzrahmen mit Maschendraht), die es uns erlauben
sollten, die neu errichteten an sich schon kleinen Quarantänekäfige (1 x 1 x 2,5 m) noch
weiter zu verkleinern. So wollten wir geschwächte Tiere vor dem Sturz auf den Käfigboden
bewahren.20
Zudem installierten wir in jedem der drei für die Quarantäne zur Verfügung stehenden
Käfige mehrere Holzleitern. Nach diesen Vorbereitungen warteten wir auf den Anruf,
wann die Tiere in Frankfurt ankommen würden. Endlich, am 14. Juli 1975, war es
soweit, die Affen sollten gegen Abend eintreffen. Ich machte mich, gemeinsam
mit meiner Frau, auf den Weg nach Frankfurt, um bei Ankunft schon am
Flughafen zu sein. Auf der Höhe von Grünberg, ca. 70 km vor Frankfurt, gab
unser Motor jedoch plötzlich auf, ich musste mich nach Grünberg abschleppen
lassen, wo festgestellt wurde, dass ein neuer Motor einzubauen sei. Daher
mieteten wir einen Leihwagen und fuhren nach Frankfurt weiter. Am Flughafen
angekommen erfuhr ich, dass die Kapuziner zwar eingetroffen, dass aber um diese
Zeit, 22.00 Uhr, sowieso kein Veterinär mehr die Tiere ansehen würde, wir
könnten die Tiere ja morgen mitnehmen. - In unserer Phantasie sahen wir
vor unseren Augen nun schon wieder Kapuzinerleichen. - Mehrere Anrufe
bei den Flughafenveterinären, „ein Verrückter bestünde darauf, die Sendung
noch heute mitzunehmen“, hatten Erfolg. Es wurde erreicht. dass einer der
Veterinäre sich die Tiere ansah und feststellte, dass alle lebten, nur waren die
Begleitpapiere, das vorgeschriebene Gesundheitszeugnis des Exportlandes,
falsch. Statt für Affen war es für Papageien ausgestellt. - Auch stimmte die
Anzahl der Tiere (11) mit der Angabe auf den Begleitpapieren (10) nicht
überein, was wir aber erst in Kassel feststellten. Die Affen durften also eigentlich
nicht eingeführt werden. Nach längeren Diskussionen hatte aber auch hier der
Flughafenveterinär ein Einsehen und erlaubte uns, die Tiere mitzunehmen.
Glücklich kehrten wir mit unserer „Last“ nach Kassel zurück. Gleich nach
Ankunft ließen wir die Tiere in die vorgefertigten Käfige. Alle lebten, waren zwar
nur teilweise adult, aber zumindest keine Säuglinge mehr. Unsere Freude
wurde auch dadurch überhaupt nicht beeinträchtigt, dass die - offensichtlich
ungeschwächten - Tiere sofort begannen, nicht nur die vorbereitete Nahrung
zu verspeisen, sondern auch die gesamte Inneneinrichtung, Leitern, Decken
und Böden, systematisch und erfolgreich zu zerstören. Sie verarbeiteten das
gesamte Fichtenholz zu großen Fasern, wobei wir sie frohgestimmt beobachteten:
Hatten wir doch unser erstes „Ergebnis“: Fichtenholz ist nicht geeignet. In der
Folgezeit verwendeten wir Buche (Stangen), Mahagonie (Kanthölzer) und Eiche
(Bretter). Hier begann nun gleichzeitig der erfolgreiche Abschnitt der Kasseler
Kapuzinerhaltung.
Durch diesen Import nun auch ausgewachsener Weibchen konnten wir auch
unsere Tiere systematisch bestimmen ([87]). Sie gehörten der Unterart Cebus
apella cay ILLIGER, 1815, an. Bubi dagegen war Vertreter einer anderen
Unterart, ihn haben wir dann der Unterart Cebus apella libidinosus SPIX, 1823,
zugeordnet.21
Darüber hinaus bescherte uns dieser
Import22
erste „Zuchterfolge“, zwei Weibchen waren schwanger und trugen ihr Junges auch aus.
Bereits am 22.08.1975 überraschte uns Teufel mit der ersten Geburt eines
Kapuzineraffen. Wir waren glücklich! Wenige Tage später schrieb ich einen Brief an
Angela Nolte und lud sie ein, nach Kassel zu kommen. Sie kam auch und
freute sich mit uns. Der kleine Kapuzineraffe wurde quer zur Längsachse
auf dem Rücken getragen und regelmäßig gesäugt. Dabei trinkt der kleine
am Bauch hängende Kapuzineraffe erst an der einen, dann an der anderen
Brust und klettert selbständig auf den Rücken zurück. Der Vorgang geht sehr
schnell, eben ist der Säugling noch auf dem Rücken der Mutter und plötzlich
am Bauch. Trotz sorgfältiger Beobachtungen hatte ich mir eingebildet, der
kleine Säugling klettere mit dem Kopf voran erst zu der einen, dann zu der
anderen Brust. Tatsächlich (den Videoaufnahmen sei Dank) hebt die Mutter
ihren Arm, das Junge lockert den Griff der Hände und lässt sich auf die eine
Brust fallen, dann wechselt es zur anderen und klettert dann auf den Rücken
zurück.
Umso trauriger waren wir, als wir unser erstes Kapuzineraffenkind am 26.09.1975 tot
vorfanden. Undenkbar war damals für uns, es könnte von Artgenossen getötet worden
sein. Als Todesursache notierte ich „Fall auf den Käfigboden“. Heute bin ich sicher,
das Kind wurde gezielt getötet, wahrscheinlich sogar von seiner Mutter. Gleich nach
der Geburt nämlich begann Teufel um Bubi zu werben, der aber nur mit Drohen
reagierte. Erst nach dem Tod des Kindes konnte Teufel enge soziale Kontakte zu Bubi
etablieren.
Über die sozialen Beziehungen der neuimportierten Tiere habe ich detailliert berichtet
([241]), auch die Fusion der zufällig zusammengesetzten drei Gruppen zur B-Gruppe
und die Fusion der A-Gruppe (Tschiggo, Purzel, Hoppediz, Bubi) mit der B-Gruppe
gelang.23
Parallel mit der Geburt des kleinen Kapuzineraffen (s. o.) nahmen die Konflikte in der
Gruppe zu, so dass wir die am meisten bedrohten Individuen separieren mussten.
([241]).
Kurzgesagt war es nicht möglich, alle Kapuziner langfristig gemeinsam in einer
Gruppe zu halten, da Teufel erfolgreich versuchte, sich die Beziehung zu Bubi exklusiv
zu sichern, indem sie nach der Geburt von Toko, dem ersten in Kassel gezeugten
Kapuzineraffen, fast alle anderen Tiere aus der Gruppe verdrängte. Durch
verschiedene Experimente und unterschiedlichste Kombinationen erhielten wir
schließlich drei Gruppen, die relativ friedlich miteinander im gleichen Raum mit
Sichtkontakt zusammenlebten. Eines der wichtigsten Resultate dieser Experimente
war, dass in jeder Gruppe ein Männchen und ein Weibchen dominierten.
Darüber hinaus war besonders auffällig, dass Individuen Partner des eigenen
Geschlechtes solchen des anderen Geschlechtes vorzogen, dass also Weibchen
bevorzugt mit Weibchen und Männchen mit Männchen interagierten. Für uns
überraschend war zudem, dass die Weibchen im Oestrus um die Männchen
warben, dass also Kopulationen von den Weibchen „hervorgerufen“ werden
müssen.24
Die Weibchen nähern sich dazu mit charakteristischer Mimik und Vokalisation
(Oestrusgesicht) einem bestimmten Männchen und versuchen, dieses zu animieren,
sie zu verfolgen. Kapuzinermännchen scheinen aber grundsätzlich wenig an
oestrischen Weibchen interessiert, so dass diese es immer wieder erneut versuchen
müssen.
Jahre später - in der neuen Primatenstation - konnte Elisabetta Visalberghi ([228]) unsere Beobachtungen
bestätigen25,
sie kam zu dem Schluss, dass das Sexualverhalten der Kapuzineraffen ein weiteres
Charakteristikum ist, um den Kapuzineraffen eine einzigartige Stellung unter den
Affen der neuen Welt zuerkennen zu müssen.

Besonders auffällig war weiterhin, dass anfänglich nur ein einziges Männchen
besonders attraktiv war. In unserem Fall war dies der alte Bubi, der uns Menschen
wenig attraktiv schien. Alle anderen - nach unserer Ansicht weit schöneren und
objektiv stärkeren - Männchen fanden wenig Beachtung. Da Bubi nun mit Teufel
separiert war, alle anderen Weibchen jedoch nur um ihn (durch das Gitter) warben,
waren die Zuchterfolge bescheiden. Sehen wir von der schon im Freiland gezeugten
Jeanette ab, bekam nur Teufel ein im Sozialverband gezeugtes Junges (Toko)). Ein
weiteres Jungtier „erzeugten“ wir, durch das kurzfristige Vergesellschaften von Ama
mit Bubi (Resultat: Kim). Heute bin ich sicher, unsere Misserfolge bei der Zucht der
Kapuzineraffen beruhten darauf, dass unsere Männchen nicht alt genug waren, um
zu züchten. Dies hatten wir freilich nicht erkannt, es war aber wohl unseren
Kapuzinerdamen bewusst. Durch verschiedene Versuche des Zusammenlassens und
Trennens erhielten wir schließlich drei Gruppen, nämlich die am stärksten bedrängten
Individuen (Erwin, Ama und Sanga (nebst Sangas Tochter Jeanette), das
„Paar“ Bubi/Teufel (nebst Teufels Sohn Toko) und Primus, der einzige Kapuziner der
B-Gruppe, der von Teufel toleriert wurde, und die restlichen neun Individuen.
Irgendwie war mein Versuch, eine große funktionierende Sozialgruppe zu bilden,
gescheitert.
Dann machte mir Werner Meinel ein nicht beabsichtigtes und nicht
von mir gewolltes „Geschenk“, er informierte mich, er habe weitere
Kapuzineraffen bestellt, die er ungefragt für eine mit Affen arbeiten wollende
Kollegin26
bestellt hatte. Ich sollte mich um die Quarantäne kümmern. Ohne einen
Quarantäneraum zu besitzen, vereinbarte ich mit den zuständigen Veterinären, alle
Kapuzineraffen unter Quarantäne zu stellen, als Quarantänekäfige sollten die
drei bereits installierten zusätzlichen Käfige (s.o.) dienen. Am Vortag der
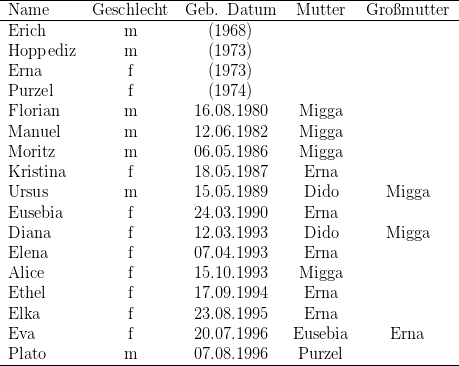
Ankunft (11. Juli 1977) bekam Teufel ihr zweites Kind, Edith, das sie tadellos
annahm. Am Abend des 12. Juli 1977 holte ich die sechs neuen Kapuzineraffen
in Frankfurt ab und überführte sie in unsere Haltung. Durch diesen Import
erhielten wir erstmals auch ein großes Männchen (Don), das uns belegte, wie ein
erwachsener Kapuzinermann auszusehen hat, ein jungadultes Männchen, Jens, drei
adulte Weibchen, Sylvia, Page und Dunka, und ein jungadultes Weibchen,
Lena. Don war für Teufel und gerade für Teufel offensichtlich attraktiver als
Bubi. Teufel sah Don, streifte ihr ein Tage altes Kind ab (Edith fiel auf den
Boden) und begann (durch das Gitter) um Don zu werben. Das schreiende
Kapuzinerbaby wurde von ihr nicht mehr beachtet. Teufel holte sogar Futter vom
Boden, direkt neben dem Kind, ohne auch nur Anstalten des Kümmerns zu
zeigen.27
Bereits in der zweiten Woche nach Import mussten Page und Dunka
- offensichtlich von Sylvia verdrängt - aus der Gruppe entfernt
werden28,
Jens und Lena hielten sich vornehmlich in einem
Fluchtkasten29
oder am Boden auf. Don und Sylvia waren deutlich dominierend. Nach Abschluss der
Quarantäne, also sechs Wochen nach Import, ließ ich einen weiteren Käfig (Käfig 7)
errichten. Die sechs neu importierten Tiere hielten wir dann gemeinsam in den Käfigen
6 und 7 des Haltungsraumes hier trennten sich die Tiere auch räumlich, Page, Dunka
und Jens hielten sich vermehrt in Käfig 6, Don, Sylvia und Lena in Käfig 7 auf.
Bubi, Teufel, Toko und Primus überführten wir in den umgebauten Käfig 5,
sie hatten also Gitterkontakt zur C-Gruppe. Beide Gruppen hatten zudem
optischen und akustischer Kontakt zu allen anderen Kapuzinern, die in Käfig 3
(Iggo-Gruppe) bzw. Käfig 4 (Erwin-Gruppe) untergebracht waren. Die beiden
dominierenden Männchen Don und Bubi bedrohten sich gegenseitig und kämpften
wenige Tage später miteinander durch das Gitter. Bubi freilich war chancenlos.
Der physischen Kondition entsprechend führte der unerbittliche Kampf bei Bubi zu schweren Verletzungen an Schwanz, Fuß und Hand, während Don außer dem Verlust einiger Haare unverletzt blieb.30
Nach diesem Kampf mied Bubi das Gitter zum benachbarten Käfig, Teufel hingegen,
die bis dahin Bubi stets stundenlang geputzt hatte, stellte die soziale Körperpflege
Bubis völlig ein und warb durch das Gitter um Don. Da dieser Zustand andauerte,
sperrten wir sie am 26. August 1977 zur C-Gruppe. Dort umwarb Teufel Don und
„erreichte“, dass dieser bereits wenige Minuten nach der Introduktion mit ihr
kopulierte. Anschließend wurde sie von Sylvia und Page bedroht. Teufel bezog das
Drohen offensichtlich nicht auf sich, drehte sich vielmehr um und drohte in die
gleiche Richtung wie Sylvia. Als Sylvia sie jedoch angriff, führte dies zu lang
andauernden Beschädigungskämpfen, die Teufel für sich entschied. Die anschließende
Introduktion von Primus und Toko verlief problemlos. In den der Introduktion
folgenden Tagen und Wochen waren drei Untergruppen zu beobachten. die
sich auch in getrennten Käfigen aufhielten, Teufel/Primus/Toko in Käfig 5,
Dunka/Page in Käfig 6 und Sylvia/Lena/Jens in Käfig 7. Don hielt sich vornehmlich
ebenfalls in Käfig 7 auf, bewegte sich aber grundsätzlich ungehindert in allen drei
Käfigen.
Den stark geschwächten Bubi introduzierten wir (ebenfalls am 26. August 1978) in die
Iggo-Gruppe.31
Hier wurde er sofort von Tschiggo, Purzel und Hoppediz begrüßt und
geputzt und suchte mühselig einen höher gelegenen Ruheplatz auf, Iggo
hingegen war vermehrt auf dem Boden zu finden und zeigte stereotypes
Kreislaufen.32

Bei erstem Gitterkontakt zwischen Don- und
Bubi-Gruppe33
(am 16. 09. 1978 in den Außenkäfigen) nahmen Teufel, Toko und Primus Kontakt zu
den Mitgliedern der Bubi-Gruppe auf. Don wurde von Iggo und Erich begrüßt.
Plötzlich fing jedoch Erich an, Angstschreie von sich zu geben, daraufhin stürzten
mit Ausnahme von Bubi, Inge, Erna und Baja alle anderen Mitglieder der
Bubi-Gruppe an das Gitter und drohten. Don wich zurück und drohte ebenfalls
aus sicherer Entfernung. Gleichzeitig drohten Teufel, Primus und Toko mit
dem Rücken zur Bubi-Gruppe ihren neuen Gruppenmitgliedern, Teufel sogar
Don.34
Auch ohne die „Unterstützung“ der Bubi-Gruppe nahmen im Oktober 1977 die agonistischen Aktionen von Primus gegenüber Sylvia, Lena und Jens zu, vor allem verstärkte Primus stets ein Drohen Teufels. Daher introduzierten wir ihn am 22. November 1977 in die Bubi-Gruppe. Sogleich nach Introduktion wurde Primus von Hoppediz, Tschiggo, Erich, Iggo, Migga und Purzel begrüßt. Neben dem üblichen Begrüßungsverhalten, wie Umarmen, Besteigen, Beriechen und Betasten der Genitalien, waren hier auch laute Begrüßungsschreie zu hören. Anschließend an die Begrüßungsphase griff Primus ohne ersichtlichen Grund Tschiggo an, worauf er von Hoppediz, Erich, Migga, Purzel und auch Tschiggo mehrfach gejagt wurde, was bei Primus zu stereotypen Kreislaufbewegungen führte. Bereits am nächsten Tag war Primus jedoch integriert und suchte besonders den Kontakt zu Iggo, dem physisch stärksten Männchen der Gruppe.
Wir stellten also fest, dass alle Individuen des A- und B-Transportes zwar nicht
gemeinsam zu halten waren, dass sie sich aber dennoch gegenseitig unterstützten bei
Anwesenheit bzw. bei Auseinandersetzungen mit den Neuimportierten. Auch diese
sechs Tiere waren nicht gemeinsam zu halten, halfen sich aber gegenseitig gegenüber
den A- und B-Tieren. Offensichtlich waren die Individuen des eigenen Transportes
ihnen vertrauter als die des anderen. Diese Situation nutzten wir aus, um eine große
Kapuzinergruppe bilden zu können ([254]).
Wir fusionierten sukzessive alle Individuen der
Don-Gruppe35 und
der Erwin-Gruppe36
- die Bubi-Gruppe (11 Tiere, Käfig 3) war für vorliegende Untersuchung ohne
Bedeutung. Die Erwin-Gruppe (Käfig 4) bestand aus dem adulten Männchen
Erwin und zwei adulten Weibchen mit jeweils einem weiblichen Jungtier,
nämlich Ama mit der am 20.08.1977 geborenen Bubi-Tochter Kim und
Sanga mit Jeanette (vgl. Tabellen 13.2 und 13.3). Ama war dominierendes
Weibchen.37
Durchgängiges Phänomen der gesamten Versuchsserie war der starke Zusammenhalt
der jeweils gemeinsam importierten Individuen. Um dies deutlich werden zu lassen,
gebe ich in diesem Abschnitt zusätzlich zu dem Namem des jeweiligen Individuums in
Klammern die Importgruppe an.
Vor Beginn der eigentlichen Experimente separierten wir am 30.11.1977 die beiden am
stärksten bedrohten Weibchen der Don-Gruppe, Dunka (C) und Page (C), in Käfig 5
und fusionierten sie mit der Erwin-Gruppe (Käfig 4). Page (C) und Dunka (C), deren
Randpositionen in der Don-Gruppe gleich nach Import (12.07.1977) auffällig
waren (s. o.), hatten bis zu diesem Zeitpunkt keinen direkten Kontakt zu
Mitgliedern der Erwin-Gruppe. Sogleich nach Fusion griffen Ama (B) und Sanga
(B), ihre Gruppe im Hintergrund, die beiden introduzierten Tiere an und
verwickelten sie in heftige Beschädigungskämpfe. Erwin (B) und Jeanette hingegen
beschränkten sich auf bloßes Drohen gegenüber Page (C) und Dunka (C). Es war also
offensichtlich nicht möglich, die Tiere der Erwin-Gruppe mit diesen beiden Weibchen
„friedlich“ zusammenzuhalten. Wir trennten daher nach zwanzig Minuten die
Erwin-Gruppe wieder ab.
Zu den in Käfig 5 separierten Weibchen Page (C) und Dunka (C) ließen wir am
01.12.1977 Sanga (B) (aus der Erwin-Gruppe) zu. Obwohl Page (C) und Dunka (C) zu
zweit und im Gegensatz zu Sanga (B) mit Käfig 5 vertraut waren, drohte Sanga (B)
nach Introduktion. Ihr Angriffsverhalten wurde durch die übrigen Mitglieder der
Erwin-Gruppe (Käfig 4) und durch Teufel (B) (Käfig 6, 7) verstärkt, nicht jedoch
durch die restlichen Tiere der Don-Gruppe. Bereits am nächsten Tag hatte sich ein
ausgewogenes Verhältnis zwischen den Gruppenmitgliedern Sanga (B), Page (C),
Dunka (C) eingestellt. Teufel (B) drohte zwar weiterhin gegen Page (C) und
Dunka (C), doch setzte Sanga (B) diese Angriffe nicht mehr fort. Page (C) und
Dunka (C) saßen jetzt häufig gemeinsam im Zwischengang oder auf einem der
Schlafkästen, während Sanga (B) sich meistens alleine und auf dem Boden
bewegte. Im Laufe der darauffolgenden Tage entwickelten sich positiv soziale
Beziehungen zwischen Page (C), Dunka (C) und Sanga (B), die sich in Kontaktsitzen
äußerten.
In die somit stabilisierte Sanga/Page/Dunka-Gruppe, in der Page (C) und Dunka (C)
dominierten, introduzierten wir am 13.12.1977 als nächstes Tier Sangas (B) Tochter
Jeanette. Kaum hatte sich der Schieber geöffnet, griff die sich am Boden befindliche
Sanga (B) sofort Page (C) und Dunka (C) an. Sie war sich offensichtlich sicher, von
ihrer Tochter unterstützt zu werden. Jeanette suchte nach der Fusion vor allem
sozialen Sitzkontakt zu ihrer Mutter, wobei auch gegenseitige soziale Körperpflege zu
beobachten war. Page (C) und Dunka (C) hingegen separierten sich und hielten sich
fast ausschließlich wieder gemeinsam im Zwischengang auf. Anfänglich drohte
Jeanette gemeinsam mit Sanga (B) gegen Page (C) und Dunka (C), vornehmlich
wieder dann, wenn Teufel (B) aus dem Nachbarkäfig drohte, doch schon am
zweiten Tag nahm sie positiv-sozialen Kontakt zu Page (C) und Dunka (C)
auf.
In diese Gruppe, in der eindeutig Sanga (B) und Jeanette dominierten, introduzierten
wir am 18.12.1977 das subadulte Männchen Jens (C) aus der Don-Gruppe, das in
dieser Gruppe häufig von Teufel (B) bedroht wurde. Sofort nach Introduktion
suchte Jens (C) den Kontakt zu Sanga (B) und Jeanette, beroch diese und
saß mit diesen im engen Körperkontakt. Zudem nahm er besonders enge
Beziehungen zu Page (C) auf. Obwohl Teufel (B) aus dem Nachbarkäfig weiter
gegen Page (C) und Dunka (C) drohte, zeigten jetzt Sanga (B) und Jeanette
agonistisches Verhalten nur noch gegenüber Dunka (C), die zusätzlich jetzt
auch von Page (C) bedroht wurde. Im Gegensatz zu dem agonistischen
Beziehungsgefüge waren positiv soziale Kontakte zwischen Sanga/Jeanette und
Page jedoch nicht mehr zu beobachten. Hingegen putzten Sanga (B) und
Dunka (C) sich gegenseitig. Jens (C) war offensichtlich attraktivstes Tier in der
Gruppe, hatte er doch zu allen Mitgliedern seiner neuen Gruppe positiv sozialen
Kontakt.38
Sofort nach Introduktion des adulten Männchens Erwin (B) am 05.01.1978 stand
dieses als dominierendes Männchen im Mittelpunkt der Gruppe, wobei Erwin (B) den
intensivsten Kontakt zu Jens (C) und Dunka (C) aufnahm, mit denen er vorher nicht
zusammengehalten wurde. Die Gruppe zerfiel offensichtlich in zwei Untergruppen,
nämlich Sanga/Jeanette und Page/Dunka/Jens, wobei besonders auffällig war,
dass Dunka nun wieder stärkeren positiv sozialen Kontakt (Kontaktsitzen,
gegenseitige soziale Körperpflege) zu Page und Jens aufnahm. Sanga und
Jeanette hielten sich nun öfter am Boden auf, hatten also offensichtlich ihre
dominierenden Positionen verloren, auch wurde jetzt Sanga (B) von Page (C)
bedroht.
Durch Erwins Wechsel in den Käfig 5 bestand für ihn nun auch Gitterkontakt zu
Mitgliedern der Don-Gruppe. Schon am Abend des ersten Tages spielte Erwin außer
mit Jens auch mit Don und Toko am Gitter. Teufel (B) hielt sich weiter am Boden auf.
Obgleich bis zu Erwins Introduktion kein gegen Teufel gerichtetes Verhalten
beobachtet werden konnte, sprachen am Abend des gleichen Tages Verletzungen bei
Teufel (B) und Lena (C) für einen Kampf zwischen beiden Tieren. Während sich in
den folgenden Tagen in der Erwin-Gruppe keine agonistischen Interaktionen feststellen
ließen, verstärkte sich das agonistische Verhalten Lenas (C) gegenüber Teufel (B),
wobei sich auch Sylvia (C) beteiligte, was zu heftigen Beschädigungskämpfen
führte.
Am 10.01.1978 introduzierten wir Teufel (B) und ihren Sohn Toko gemeinsam in die
Erwin-Gruppe. Sogleich nach Introduktion fiel auf, dass Teufel, die sich in der
Don-Gruppe zuletzt nur am Boden aufgehalten hatte, in der Erwin-Gruppe sofort in
die oberen Käfigbereiche kletterte. Während die neuen Gruppenmitglieder Teufel sehr
„vorsichtig“ behandelten - kein anderes Tier der Gruppe nahm Interaktionen zu
Teufel auf - wurde Toko sogleich von beiden Männchen, Erwin (B) und Jens (C),
„begrüßt“. Anschließend bestieg Toko mehrfach Erwin und Jens und versuchte auch
bei Sanga (B) aufzureiten. Nach zwei Tagen hatten Teufel (B) und Erwin (B) positiv
soziale Beziehungen zueinander aufgenommen, erkennbar an sozialer Körperpflege und
Kontaktsitzen, wobei die Interaktionen meist von Erwin ausgingen. Toko spielte
weiterhin mit Jens, Erwin nur mit Toko. Während Sanga (B) in dieser Gruppe
isoliert war, nahm Jeanette zu Erwin, Teufel und Toko Kontakt auf. Page (C)
und Dunka (C) wurden von Teufel (B) und Toko (Page) bzw. von Sanga
(B) (Dunka) bedroht und nahmen Randpositionen in der Gruppe ein (Abb.
5).
Am 14.01.1978 fiel auf, dass Teufel, obwohl seit geraumer Zeit gravid, Erwin, mit dem
sie nun vier Tage in einer Gruppe zusammengehalten wurde, mit hochgezogener Stirn
und horizontal nach hinten gezogenen Mundwinkeln ansah (Oestrusgesicht). Sie stieg
anschließend zu Erwin auf den Kasten und legte sich vor dieses Männchen, worauf
Erwin auf Teufel aufritt. Dieses Verhalten konnte jedoch nur einmal beobachtet werden
(s. u.).
Als letztes Tier der ehemaligen Erwin-Gruppe introduzierten wir die seit dem
05.01.1978 allein mit ihrer Tochter Kim gehaltene Ama (B) am 15.01.1978 zur
Erwin-Gruppe, der damit zusätzlich Käfig 4 gegeben werden konnte. Gleichzeitig
bekamen hierdurch alle Mitglieder dieser Gruppe auch die Möglichkeit des
Gitterkontaktes zur Bubi-Gruppe. Nach Öffnen des Verbindungsganges zwischen den
Käfigen 4 und 5 lief Ama (B) sofort zur Erwin-Gruppe und nahm dort schon nach
kurzer Zeit sozialen Sitzkontakt zu Teufel (B) auf. Toko dagegen lief in den Käfig 4 an
das Gitter zur Bubi-Gruppe, wo er von mehreren Mitgliedern dieser Gruppe
„begrüßt“ wurde. Dabei umarmten sich die Tiere und betasteten gegenseitig
ihre Genitalien. Zugleich wurden laute Begrüßungsschreie ausgestoßen. Nach
dieser Begrüßung lief Toko zu dem neu introduzierten Weibchen Ama (B),
betastete es und versuchte aufzureiten. Im Folgenden nahm Ama (B) dann auch
positiven Kontakt zu Sanga (B) auf. Sanga saß dabei häufig neben Ama
und ließ sich von Ama putzen. Der positive Kontakt von Erwin (B) und
Jens (C) zu Toko bzw. von Erwin zu Teufel ließ nach Amas Introduktion
nach, auch nahmen Erwin (B) und Jens (C) keinen weiteren Sozialkontakt zu
anderen Tieren auf, wodurch sie eine isolierte Position innerhalb der Gruppe
einnahmen.39
Bereits am Abend des 16. 01. 1978 konnten wir beobachten, wie Kim alleine
in der neuen Gruppe umherkletterte. Da nach unseren Beobachtungen alle
Jungtiere in Gefahrensituationen auf dem Rücken ihrer Mütter bleiben bzw. zu
diesen flüchten, musste wohl schon hier eine eindeutige Klärung stattgefunden
haben.
Da die Erwin-Gruppe nach Introduktion von Ama vor dem Auseinanderfallen
war, wir hätten alle drei Individuen des C-Importes, Page, Dunka und Jens,
isolieren müssen, introduzierten wir bereits am 17.01.1978 das Weibchen Lena
(C) aus der Don-Gruppe. Die erste Zeit nach der Introduktion hing Lena
fast ausschließlich am Verbindungsgitter zu Käfig 6. Nach etwa 15 Minuten
bewegte sie sich auch vorsichtig auf dem Klettergerüst in Käfig 5, floh anfangs
jedoch, wenn ein anderes Mitglied der Erwin-Gruppe sich näherte, sofort
an das Gitter zu Don und Sylvia. Allmählich blieb Lena (C) bei Jens (C)
Annäherungen auch sitzen und nahm nach etwa einer halben Stunde den ersten
Sozialkontakt zu diesem Männchen auf, der sich allerdings auf Kontaktsitzen
beschränkte. Über Jens schloss sich Lena (C) auch Page (C) und Dunka (C)
an.40
In den der Introduktion folgenden Tagen verringerten sich Lenas (C) soziale
Beziehungen zu Page (C) und Dunka (C), sie nahm nun Kontakt zu Teufel (B) und
Erwin (B) auf. Während Page (C) zunehmend häufiger sozialen Kontakt zu Ama
(B), Teufel (B) und Sanga (B) unterhielt, blieb Dunka (C) innerhalb der
Gruppe isoliert und wurde auch besonders häufig von Ama (B) und Teufel (B)
bedroht.
Am 22.01.1978 introduzierten wir Sylvia (C) in die Erwin-Gruppe. Zunächst ergaben
sich keine Gruppenveränderungen. Sylvia nahm sogleich zu fast allen Tieren, mit
Ausnahme von Dunka (C), Page (C) und Erwin (B), sozialen Sitzkontakt auf.
Agonistisches Verhalten konnte während der ersten Stunden nach Sylvias Introduktion
nicht beobachtet werden. Zwei Tage nach Sylvias Introduktion zeigte sich jedoch
erneut eine Tendenz zur Untergruppenbildung. Sylvia (C) schloss sich mehr und mehr
Lena (C) und Jens (C) an; dementsprechend ließen auch die anfangs beobachteten
Interaktionen zu anderen Gruppenmitgliedern nach. Weiterhin reduzierte Page (C)
wieder ihre Kontakte zu Sanga (B), Teufel (B) und Ama (B) und hatte erneut
häufigen Sitzkontakt mit Dunka (C). Die Spaltung der Gruppe war auch im
agonistischen Bereich deutlich. Lena, Jens, Sylvia, Page und Dunka und somit alle
Tiere der sogenannten C-Gruppe (vgl. Tabelle 13.2) wurden verstärkt bedroht
und zwar vornehmlich von Teufel, Ama und Sanga, somit von allen adulten
Weibchen der B-Gruppe. Im Folgenden suchten und fanden jedoch Dunka (C) und
Page (C) Anschluss an die Weibchen der B-Gruppe, während Sylvia (C),
Lena (C) und Jens (C) eine eigene Untergruppe bildeten, wobei nur Toko
verstärkt Spielkontakt zu Jens unterhielt. Dementsprechend waren nun auch keine
agonistischen Interaktionen mehr zwischen Sanga/Ama/Jeanette und Dunka/Page
zu beobachten. Allein Teufel drohte, wenn auch selten, als dominierendes
Weibchen allen anderen Gruppenmitgliedern. Vergleichbar mit dem dominierenden
Männchen Erwin hatte sie nur noch wenige Kontakte zu anderen Mitgliedern der
Gruppe.
Den Abschluss der Integration beider Gruppen bildete am 07.02.1978 die Introduktion
des seit zwei Wochen solitär gehaltenen Männchens Don. Da mit Verletzungen bei
Gitterkämpfen unter den Männchen Bubi und Don zu rechnen war (s. o.), sollte der
herrschende Gitterkontakt beider Gruppen noch vor der Introduktion unterbrochen
werden. Deshalb setzten wir die Bubi-Gruppe in die Käfige 5, 6 und 7 um, während
die Erwin-Gruppe mit dem zu introduzierenden Männchen Don die Käfige 3 und 4
erhielt.
Don drohte in der neuen Gruppe als erstes ungerichtet gegen die Bubi-Gruppe, zu
ihm gesellte sich Toko, der in die gleiche Richtung drohte. Anschließend war
auffallend, dass Don von allen neuen Gruppenmitgliedern - auch Toko - gemeinsam
bedroht wurde. Diese saßen hierbei im Halbkreis um den in einer Käfigecke
sitzenden Don herum. Don drohte daraufhin einmal zurück, beendete so die
gegen ihn gerichteten agonistischen Aktivitäten und übernahm gleichzeitig
unangefochten die Rolle des dominierenden Männchens in dieser Gruppe. Sylvia und
Lena suchten danach sogleich Kontakt zu Don und wurden mehrfach von ihm
geputzt.
Auffällig war auch eine Änderung im Verhalten Erwins. Hatte er vorher als
dominierendes Männchen eine isolierte Stellung in der Gruppe, so nahm er nach Dons
Introduktion zu Teufel, Ama, Sanga und Jeanette sozialen Sitzkontakt auf. Auch
spielte Erwin nun häufig mit den beiden Männchen Toko und Jens, Don hingegen nur
mit Toko (Abb. 9). In der neu gebildeten Gruppe waren agonistische Interaktionen
nicht mehr zu beobachten. Nach den positiv-sozialen Beziehungen am Tag der
Introduktion war eine Aufspaltung der Gruppe auffällig, nämlich (1) Page (C)/Dunka
(C), die völlig isoliert in der Gruppe waren und nur zu Don Kontakt hielten. (2)
Don/Lena/Sylvia und (3) der Rest der Gruppe, also alle Tiere der ursprünglichen
B-Gruppe und Jens (C), der durch Spielkontakte zu Toko und Erwin Anschluss an
diese Untergruppe fand. Diese Gruppenkonstellation blieb jedoch nur einen Tag
erhalten, bekam doch Sylvia in der Nacht nach Introduktion ein weibliches Jungtier,
Pan (vgl. Tabelle 13.3). Sie nahm nun verstärkt Kontakte zu Ama, Sanga und
Jeanette auf, wodurch sich ihre Beziehungen zu Lena und Don verringerten. Am
26.02.1978 wurde in der Gruppe noch die Teufel-Tochter Isis (vgl. Tabelle 13.3)
geboren.
In die Don-Gruppe integrierten wir zudem ab dem 16.03.1978 die handaufgezogene Teufel-Tochter
Edith41
und im Laufe des Jahres 1978 sukzessive Erna (mit ihrer Tochter Claudia),
Iggo, Purzel, Baja und Tschiggo. So erreichten wir durch diese Experimente
zwei mehr oder weniger stabile Gruppen. Die Don-Gruppe bestand Ende
1978 aus 22 Individuen, 16 Wildfängen und sechs bei uns geborenen
Jungtieren. In beiden Gruppen wurden weitere Jungtiere geboren (vgl. Tabelle
13.3)42
Unsere Erfolge (s. o.) bestätigten im Nachhinein unser Vorgehen. Bevor wir auf einige
Resultate näher eingehen, sei nur betont, dass diese Gruppen bis zum Ende der
Kapuzinerhaltung (1996) bestanden und dass über die Jahre die Präferenz für
Individuen des selben Importes - wenn auch nicht mehr so deutlich - aufzeigbar
waren.
In der Bubi-Gruppe, die dann versuchsbedingt und ohne Zwang mehrfach
verkleinert wurde, blieb Bubi unangefochten bis 1982 dominierendes Männchen.
Dann übernahm Erich diese Position.43 Erichs Ägide war aber nur kurz, ein Jahr später verlor er nach einem harten Kampf mit dem zehnjährigen Hoppediz die Position des dominierenden Männchens, Erich musste separiert werden. Die ehemalige Bubi-Gruppe wurde zur Hoppediz-Gruppe. Da unsere Untersuchungen gezeigt hatten, dass freundschaftliche Beziehungen unter Kapuzineraffen stabil sind und Bubi und Hoppediz stets freundschaftliche Beziehungen unterhalten hatten, ging ich das Risiko ein, Bubi in seine ehemalige Gruppe zu reintroduzieren. Das Experiment hatte aber wohl dramatische Konsequenzen für die in der Gruppe geborenen Kinder, worüber ich noch berichten werde, und auch für mich.44
Nach unseren Daten werden Kapuzinerweibchen das erste Mal im Alter von 3,8 J ± 0,4 Jahren oestrisch und werben um Männchen der Gruppe. Die ersten Geburten erfolgten im fünften Lebensjahr (s. o.). Grundsätzlich können wir, da wir unsere Affen nur beobachten, über den genauen Zeitpunkt der hormonell gesteuerten Oestren keine Auskunft geben. Unsere Daten beschränken sich stets auf den

„Verhaltensoestrus“, also auf das protokollierte Werben mit anschließender Kopulation
bzw. auch auf permanentes erfolgloses Werben. Hier haben wir unzählige Male diesen
Verhaltenskomplex protokolliert. Einschränkend muss betont werden, dass
offensichtlich (zumindest drei Fälle) auch gravide Weibchen oestrisch werden können,
um Männchen werben und auch bestiegen werden (mit Intromissio). Diese
„Scheinoestren“ sind in unserem Datenmaterial auch enthalten und können nach dem
Verhalten von echten Oestren nicht diskriminiert werden. Nach diesen Daten dauert
die Zeitspanne von Beginn des letzten Oestruses vor der Gravidität bis zur
Geburt 157 ± 4 Tage und beträgt somit die Tragzeit mehr als 150 Tage.
Werden die Weibchen nicht gravid, dann folgt sehr bald ein neuer Oestrus
(Kapuzinerweibchen sind also polyoestral). Der Abstand der einzelnen Oestruszyklen
betrug in 44% der Fälle im Mittel 19,7 Tage, zu 45% jedoch länger (15% im
Mittel 32,1 , 10% 45,4, 15% zwischen 50 und 100 Tagen und in 5% sogar
länger als 150 Tage). Zudem folgten 11% der Oestren bereits nach 15 und
weniger Tagen (im Mittel 11,6 Tage), wobei wir vermuten, dass es sich bei
den letzten um Scheinoestren handelt. Die meisten Oestren beobachteten
wir im Januar und Februar, die wenigsten in der Zeitspanne von Juli bis
Oktober.
Sabine Schulz berichtet in ihrer Examensarbeit: „Während unserer Versuche
beobachteten wir eine allgemeine Zunahme des agonischen Verhaltens aller Tiere
während und nach dem Oestrus eines oder mehrerer Weibchen. Schon während der
Zeit des Oestrus eines Weibchens nimmt das agonistische Verhalten unter den
anderen Gruppenmitgliedern zu. Bemerkenswert ist dabei, dass das oestrische
Weibchen nahezu niemals bedroht wird. Dies könnte dadurch erklärt werden,
dass ein Bedrohen des oestrischen Weibchens ein Bedrohtwerden von seiten
des umworbenen Männchens nach sich ziehen könnte. Dies ist offensichtlich
unabhängig davon, ob das Männchen auf das Werben des Weibchens antwortet oder
nicht.
In der Don-Gruppe wurde z. B. regelmäßig beobachtet, dass das dominante Weibchen
Sylvia versuchte, oestrische Weibchen davon abzuhalten, sich dem dominanten
Männchen Don zu nähern. So näherte Sylvia sich immer wieder Don, woraufhin das
oestrische Weibchen vor ihr wich. Ging Don auf das Werben ein, d. h. zeigte er die
gleichen Verhaltensweisen (Oestrusgesicht, charakteristische Lautäußerung),
„floh“ das Weibchen vor ihm, um sich verfolgen zu lassen, was dann in eine
Kopulation einmünden konnte. Diese „Flucht“ und das „Verfolgen“ scheint
obligatorisch zu sein und ist dasjenige Verhalten, das das Weibchen zu initiieren
versucht.

... Weiterhin stellten wir während unserer Beobachtungen fest, dass grundsätzlich oestrische Weibchen ihre Sozialkontakte zu anderen Tieren reduzieren. Das oestrische Weibchen hat keine Kontakte mehr zu irgendeinem Tier der Gruppe, es zeigt vielmehr nur noch Interesse an dem jeweils umworbenen Männchen. Oestrische Weibchen zeigen daher weder positiv soziale noch agonistische Verhaltensweisen“ (Seite 78-79). Selbst eigentlich eng befreundete Weibchen halten während der Oestrusphasen keine Sozialkontakte untereinander, hierfür gibt Sabine Schulz mehrere Beispiele.

Sabine Schulz berichtete auch kurzfristige Veränderungen des sozialen Beziehungsgeflechtes bei Geburten: „Grundsätzlich konnten wir beobachten, dass Mütter für alle Gruppenmitglieder attraktiv sind. Hierdurch wird verständlich, dass sich die soziale Position eines Weibchens durch die Geburt seines Kindes verbessert. Mütter haben zu Müttern in der Regel intensive Beziehungen, auch wenn sie vor den Geburten ihrer Kinder nur geringfügige (Kontakte) zueinander hatten. Waren die Beziehungen zwischen zwei Müttern schon vor den Geburten eng, so werden sie nach den Geburten noch intensiver. Während die Kontakte zwischen den Müttern erheblich zunehmen, werden die Beziehungen zu allen anderen Weibchen, zu denen die Mütter vorher Kontakte hatten, von den Müttern erheblich, oder zumindestens relativ im Vergleich zu den Beziehungen zu anderen Müttern, reduziert. Wenn die Kinder etwas größer sind und nicht mehr ständig getragen werden, werden die Beziehungen zwischen den Müttern wieder zunehmend geringer, bis sie wieder die Stärke erreicht haben, die schon vor den Geburten beobachtet wurde“ (Seite 80). Diese Ergebnisse belegt Sabine Schulz mit zahlreichen detaillierten Beispielen.
Im zweiten Teil dieses Kapitels werde ich detailliertere Angaben zur
Entwicklung der Jungtiere angeben, hier also nur die Befunde, die wir bei
den Kapuzineraffen-Beobachtungen nebenbei erhoben haben. Neugeborene
Kapuziner sind völlig hilflos und halten sich in den ersten drei Lebenswochen mit
Ausnahme kurzer Saugzeiten überwiegend mehr oder weniger bewegungslos
auf dem Rücken der Mutter auf (dabei quer zur Längsrichtung der Mutter
orientiert). Im ersten Lebensmonat verlassen sie ihre Mütter nicht. Die einzigen
Aktivitäten, die wir beobachten können, sind Saugen, Umherschauen, Sichkratzen,
Klettern auf der Mutter und Aufrichten auf der Mutter (ab dem Alter von
zwei Wochen). Unabhängig hiervon klettern sie jedoch - wenngleich äußerst
unbeholfen - bereits im Alter von 22 Tagen (bei zwei Jungtieren in unserer Kolonie
beobachtet) auch auf den Rücken von gemeinsam mit der Mutter ruhenden
Artgenossen. Sie werden aber von diesen zu diesem Zeitpunkt noch nicht
getragen.45
Ab dem zweiten Lebensmonat - die Jungen werden nun überwiegend längs zur
Körperlängsachse getragen und sind auch lokomotorisch aktiver (lösen sich auch schon
häufig mit einer Extremität von der Mutter, um zu manipulieren) - können wir
regelmäßig das Tragen durch andere Tiere (ältere Geschwister, Weibchen und
Männchen) beobachten, wobei es sich jedoch stets um ein mit der jeweiligen Mutter
„befreundetes“ Individuum handelt. Im Alter von 29 bis 57 Tagen sitzen die Jungen
erstmals neben der Mutter (dabei engen Körperkontakt haltend). Ein Alleinsein
beobachteten wir zwischen dem 34. und 53. Lebenstag zum ersten Mal, zwischen dem
51. und 69. Tag dann regelmäßig. Kontakt zu anderen Artgenossen nehmen die
Jungtiere aktiv zwischen dem 58. und 87. Lebenstag auf. Das selbständige
Klettern und die Aufnahme festen Futters beobachten wir erstmals im zweiten
Lebensmonat. Der enge Kontakt zur Mutter dauert unabhängig von der zunehmenden
lokomotorischen Reifung bis zum Ende des sechsten Lebensmonates (zum
Schlafen und bei Gefahr wird der Rücken der Mutter aufgesucht). Darüber
hinaus werden die Jungen auch noch im elften Lebensmonat gelegentlich
gesäugt.
Betrachten wir nun die sozialen Interaktionen der Jungtiere zu anderen
Gruppenmitgliedern, dann sind, bedingt durch deren Unbeholfenheit und
ständiges Angewiesensein auf die Mutter, erwartungsgemäß die Kapuzinerkinder
anfänglich nur „Empfänger“ sozialer Interaktionen. In den ersten Lebensmonaten
sind für den kleinen Kapuziner die passiven Kontakte zu seinen Geschwistern
besonders auffällig. Anfänglich - im ersten Lebensmonat - sind an ihm vor allem
nächstältere Geschwister - unabhängig von deren Geschlecht - und ältere Schwestern
interessiert. Kontakte zu älteren Brüdern sind erst ab dem zweiten Lebensmonat
bemerkenswert.46
Hier werde ich noch speziellere Befunde vorstellen (s. u.).
Im dritten Lebensmonat suchen nun die Jungtiere selber den Kontakt zu
Geschwistern.47 Im Alter
von vier Monaten48
werden sie attraktiv für Individuen des gleichen Jahrganges, deren Nähe sie dann im
fünften Lebensmonat aktiv suchen. Ab diesem Zeitpunkt sind auch verstärkt von
adulten Männchen ausgehende Sozialkontakte zu beobachten. Hier ist es in der Regel
ein ganz bestimmtes Männchen, dass zu dem jeweiligen Jungtier Kontakt
aufnimmt. Dieses Männchen ist dann auch der nach unseren Beobachtungen
mögliche Vater, den wir - da Vaterschaftsausschlüsse bisher noch nicht geleistet
wurden - vorerst als „psychischen Vater“ bezeichnet haben. Gerade zu diesem
Männchen nimmt dann das entsprechende Jungtier im sechsten Lebensmonat
selbst Kontakt auf. Gleichzeitig ist in diesem Lebensmonat erstmals auch zu
beobachten, dass die Jungtiere neue Sozialpartner wählen, suchen sie doch
aktiv den Kontakt zu einem Jahr älteren - nichtverwandten - Juvenilen (s.
u.).
Zur Sozialstruktur ist besonders bemerkenswert, dass bei Kapuzinergruppen
Rangordnungen - wie wir sie von vielen catarrhinen Spezies kennen - als
Regulationsmechanismen des Gruppenzusammenhaltes nicht beobachtet werden
können. Die Gruppe beruht hier ausschließlich auf der Ausgewogenheit der sozialen
Beziehungen.
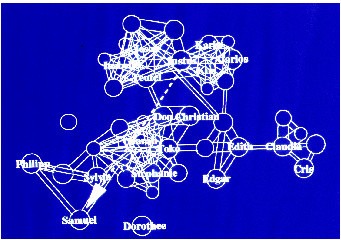
Für das komplexe Netzwerk der sozialen Beziehungen konnten zehn „Regeln“ angegeben
werden, die sich in jeder möglichen Gruppenkonstellation wiederfinden lassen.
Regel 1: In jeder Kapuzinergruppe gibt es ein dominierendes Männchen und ein
dominierendes Weibchen.
Regel 2: Das dominierende Männchen isoliert sich aktiv von dem Rest der Gruppe.
Unabhängig hiervon ist es das attraktivste Tier für alle anderen Gruppenmitglieder.
Regel 3: Das dominierende Weibchen zeigt ebenfalls Tendenzen, sich von den übrigen
Tieren zu separieren. Es versucht, besondere Beziehungen zu dem dominierenden
Männchen aufzubauen und alle anderen Tiere, die sich dem dominierenden Männchen
nähern, zu vertreiben.
Gerade die ersten drei Regeln gewannen wir bei den Gruppenbildungsexperimenten,
bei Beobachtungen an Gruppen mit unterschiedlicher Zusammensetzung der
Beteiligten und dementsprechend häufig zu beobachtenden Änderungen der sozialen
Positionen einzelner Gruppenmitglieder (s. o.). Bei dem möglichen Verlust der
dominierenden Position änderte sich nämlich auch das übrige Verhalten des
Individuums. Männchen, die vorher mehr oder weniger außerhalb der Gruppe standen,
waren nun wieder an dem Geschehen innerhalb der Gruppe beteiligt. Auch der
umgekehrte Prozess, dass Männchen plötzlich dominierend werden, konnte mehrfach
beobachtet werden.
Das dominierende Kapuzinermännchen kontrolliert die Gruppe nun aber nicht - wie
ein Alpha-Männchen der Makaken (vgl. Kapitel 14), das alle Interaktionen
beeinflusst und bei Gruppenauseinandersetzungen interveniert - es ist vielmehr ein
Orientierungspol für die anderen ohne großen Einfluss auf das Gruppengeschehen. Ihm
kommen sicherlich Aufgaben bei der Verteidigung der Gruppe gegenüber Individuen
der gleichen Art zu (und in Menschenobhut auch gegenüber dem menschlichen
Beobachter). Insofern sitzen auch die jeweiligen dominierenden Männchen in
der Peripherie der Gruppe und orientieren sich an den anderen Gruppen,
deren Interaktionen sie „kommentieren“. Entscheidend für die dominierende
Position ist offensichtlich nicht, so lehrte uns Bubi, die physische Stärke,
vielmehr ist von hoher Wichtigkeit das „Sich-dominierend-fühlen“ und das
„Als-dominierend-angesehen-werden“. Dominierende Männchen bewegen sich ohne
Furcht vor anderen, weichen bei keiner Gefahr und reagieren nicht auf das Bedrohen
durch andere.
Das dominierende Weibchen hat keine so exponierte Position wie das dominierende
Männchen. Es ist das Weibchen, das alle Auseinandersetzungen für sich entscheidet.
Ganz im Gegensatz zu der hohen Attraktivität des dominierenden Männchens meiden
alle anderen Tiere seine Nähe. Einige Beobachtungen sprechen dafür, dass man auch
im weiblichen Geschlecht, um dominierend zu sein, sich dominierend fühlen muss, wie
ich oben ausgeführt habe.
Jedes dominierende Weibchen versucht, sich das dominierende Männchen exklusiv zu
sichern, so intervenieren dominierende Weibchen bei Annäherung oestrischer
Weibchen. Erst wenn das oestrische Weibchen es geschafft hat, das dominierende
Männchen zum Nachfolgen zu bewegen, bleiben die Interventionen unbeachtet bzw.
werden eingestellt. Wir haben den Eindruck. dass dominierende Weibchen versuchen,
eine Paarbeziehung aufzubauen, was ihnen aber - wegen des Desinteresses des
dominierenden Männchens - nicht gelingt.
Regel 4: Der Rest der Gruppe gliedert sich in Untergruppen. Eine Untergruppe basiert
auf Zweier- oder Verwandtschaftsbeziebungen, wobei jedes Tier mehr als eine
Zweierbeziehung eingehen kann.
Regel 5: Zweierbeziebungen und Untergruppenbeziehungen sind sehr stabil und bleiben
erhalten, auch wenn sich die Gruppenzusammensetzung ändert.
Anfänglich konnten wir in unserer Kolonie nur Untergruppen beobachten, die auf
Zweierbeziehungen beruhten. Diese waren äußerst stabil. Geht ein Individuum
mehrere Zweierbeziehungen zur gleichen Zeit ein, dann entstehen größere
Untergruppen. Dabei ist es nicht zwingend, dass dann auch alle Tiere miteinander
interagieren. Eventuell ist eines der Individuen so attraktiv, dass es mehrere Partner
über Zweierbeziehungen bindet. Wir konnten in der Vergangenheit mehrere
Typen solcher Beziehungen beobachten, so können Zweierbeziehungen auf
der gleichen sozialen Situation (gemeinsames Verfolgen bzw. gemeinsames
Verfolgtwerden) beruhen oder aber auf „Freundschaften“. Nur die letztgenannten
Zweierbeziehungen sind langfristig stabil. Untergruppenbeziehungen beruhen auf den
sozialen Interaktionen der Weibchen. Männchen mit grundsätzlich weit lockerer
Untergruppenbindung (s. u.) zeigen gegenüber den Untergruppenpartnern das
nämliche Verhalten wie Weibchen untereinander: Kontaktsitzen und gegenseitige
Körperpflege.
Regel 6: Wird eine Kapuzinergruppe durch fremde Tiere vergrößert, können zwei
verschiedene Untergruppen gemeinsam eine neue Untergruppe bilden.
Unsere Experimente während dieser Jahre haben mehrfach gezeigt, dass es
relativ leicht möglich ist, in bestehende Gruppen neue Tiere zu introduzieren.
Kapuzinergruppen sind also weit weniger geschlossen als z. B. Makakengruppen. Bei
letzteren könnte der Ausgang (der Neuankömmling wird massiv bedroht und
angegriffen, vgl. Kapitel 14) vorhergesagt werden, bei den Kapuzineraffen
jedoch sind Prognosen grundsätzlich nicht möglich. Generell konnten wir
drei verschiedene Konsequenzen einer Introduktion beobachten. Entweder
wird der Neuankömmling (1) gleich massiv bedroht (und muss dann wieder
separiert werden) oder aber er kann mehr oder weniger unbedroht in der Gruppe
bleiben. Findet er nun (2) dort Anschluss an eine Untergruppe, die „nichts
zu sagen hat“, dann hat die Introduktion keinen Einfluss auf die generelle
Struktur des Sozialgefüges. Er kann aber auch (3) selber dominierend werden,
Anschluss an eine dominierende Untergruppe finden (diese dadurch zu sehr
verstärkend) oder aber auch durch seinen Anschluss eine vorher nicht dominierende
Untergruppe zu der oder zu einer der dominierenden werden lassen, dann hat seine
Introduktion dramatische Folgen, die zur völligen Umorientierung der Gruppe
führen können. Vorher bedrohte periphere Individuen drohen nun selber und
umgekehrt.
Regel 7: Im Gruppenalltag präferieren adulte Männchen den Kontakt mit adulten
Männchen, adulte Weibchen denjenigen mit adulten Weibchen und Juvenile -
unabhängig vom Geschlecht - denjenigen mit Juvenilen und auch mit adulten
Männchen.
Regel 8: Sozialkontakte unter Männchen und unter Juvenilen bzw. zwischen Männchen
und Juvenilen sind vornehmlich Spielkontakte. Dabei spielen Männchen and Juvenile
auch mit Männchen und Juvenilen anderer Untergruppen. Kampfspiele zwischen
Männcben beruhen auf Zweierbeziehungen, wobei ein Tier mit zwei anderen Tieren zur
gleichen Zeit spielen kann.
Noch in der provisorischen Kapuzinerhaltung mussten wir erkennen, dass die Regel 7
in der hier formulierten Form nicht mehr uneingeschränkt gilt, so lernten wir,
dass alle Männchen unserer damaligen Kolonie - mit Ausnahme von Bubi
und Don - jungadult waren. Volladulte Männchen separieren sich generell
von anderen Volladulten und neigen dazu, relativ wenig untereinander zu
interagieren.49
Sie bleiben aber für jungadulte und juvenile Männchen von höchster Attraktivität.
Ansonsten gelten beide Regeln weiterhin. Gerade beim Kampfspiel, der vornehmlichen
Aktivität, finden wir wiederum, wie bei den Untergruppenbeziehungen, die
Beschränkung auf die Dyade. Dabei können wir auch beobachten, wie ein Tier
gleichzeitig mit mehreren anderen spielt, es kämpft den einen Spielpartner nieder,
wendet sich dem zweiten zu und umgekehrt. Zeigen nun zwei Tiere das nämliche
Verhalten, können auch größere Spielgruppen entstehen, von vier bis sechs Individuen.
Neben dieser Spielform finden wir auch den sogenannten „Arena-Kampf“, den gerade
Don manchmal in den Abendstunden zeigt, dabei hüpft er biped mit ausgestreckten
Armen am Boden herum und wird von nahezu allen Juvenilen angegriffen
bzw. greift diese auch selber an. Wenn er dann, ein schweres ausgewachsenen
Kapuzinermännchen, auf ein z. B. einjähriges Jungtier hüpft, mehrfach auf
diesem auf- und abspringt, gewinnt der Beobachter leicht den Eindruck, dass
dieses malträtiert wird. Der junge Kapuziner scheint dies hingegen anders
zu empfinden; mühsam Don entkommen, springt er diesen erneut an, um
entsprechendes wieder zu initiieren. Der abendliche „Arena-Kampf“ mit teilweise
mehr als zehn Beteiligten ist das Eindrucksvollste, was unsere Kapuziner
demonstrieren.
Regel 9: Die sozialen Kontakte der Weibchen sind vor allem Kontaktsitzen mit
anderen Weibcben und gegenseitige soziale Körperpflege. Soziale Körperpflege zwischen
Weibchen verschiedener Untergruppen sind sehr selten.
Regel 10: Kapuziner verteidigen die Mitglieder ihrer eigenen Gruppe oder Untergruppe
gegenüber Fremden. Dabei ist ein Gruppenbewusstsein aufzeigbar.
Diese Leistung der Kapuziner ist auch - wie schon oben angegeben - die
Voraussetzung für die Bildung von größeren Gruppen gewesen. Die Kapuziner sind sich
ihrer komplexen Struktur offensichtlich bewusst. Bei Konfrontation mit einem
gemeinsamen Feind (eine fremde bedrohliche Person, ein Hund etc.) wird dieser von
allen Gruppen gleichzeitig bedroht, ansonsten bedrohen sich die Gruppen bzw. die
Untergruppen einer Gruppe untereinander. Bedingt durch das gleichzeitige
Halten mehrerer Gruppen in einem Raum, also durch die Anwesenheit der
Gruppenfeinde, wird in Kassel das Auseinanderfallen der Gruppen verhindert, dieser
Prozess dürfte vergleichbar mit Freilandbedingungen sein. Ohne äußere Feinde
würde nach unserer Einschätzung jegliche Gruppenkonstellation langfristig
zum Paar bzw. zur Kleinfamilie führen, da Weibchen sich dann gegenseitig
verdrängten.
Auch zur sozialen Stellung der Jungtiere werde ich noch im zweiten Teil detailliertere
Angaben leisten. Da bei Kapuzinern, wie schon betont, Rangordnungsstrukturen nicht
zu finden sind, jedes Kapuzinerkind aber eine bestimmte soziale Stellung erhält,
interessierte uns besonders, welche Einflüsse auf das in der Gruppe geborene Jungtier
einwirken bzw. welche von diesem selber gegen andere Gruppenmitglieder gerichtet
sind. Zur Lösung dieses Fragenkomplexes beobachteten wir seit 1981 im Rahmen einer
von der Deutschen Forschungsgemeinschaft geförderten Längsschnittstudie zum
Sozialverhalten von Cebus apella neben der Gruppe nun auch bestimmte
Einzeltiere und protokollierten genauestens alle ihre Interaktionen mit den übrigen
Gruppenmitgliedern. Durch das tägliche kontinuierliche Beobachten von Individuen
beiderlei Geschlechts, beginnend mit der Geburt bis zum Adultwerden, konnten wir die
Parameter bestimmen, die für die spätere soziale Stellung der Jungtiere verantwortlich
sind (s. u.).
Nach den bisherigen Beobachtungen und den Daten zur frühen postnatalen
Entwicklung (s o.) konnten einige Befunde als gesichert gelten. So interagierten -
unabhängig von Alter und Geschlecht - alle in der Gruppe geborenen Jungtiere mit
ihren Müttern und Geschwistern, wobei die Anzahl der sozialen Kontakte zu
einigen oder allen Verwandten die zu anderen Mitgliedern der Gruppe übertraf.
Darüber hinaus bevorzugten grundsätzlich alle Juvenilen den Kontakt zu
Tieren des gleichen Altersbereiches, interagierten also verstärkt mit gleichalten,
ein Jahr älteren und ein Jahr jüngeren Individuen. Entsprechend nahmen
auch die Interaktionen zu Geschwistern mit zunehmendem Altersabstand
ab.
Kontakte zu nichtverwandten Adulten scheinen nach ihrem quantitativen Auftreten
generell nicht bemerkenswert, interagieren doch Juvenile aller Altersstufen nahezu
ausschließlich nur mit Kontaktpartnern der Mutter, wobei sie hier - während
der ersten Lebensjahre - passive Partner der jeweiligen Zweierbeziehungen
sind. Mit zunehmendem Alter jedoch (ab dem dritten Lebensjahr) können die
entsprechenden Kontakte (vor allem Sitzkontakte und soziale Körperpflege, also
Verhaltensweisen, die - wie schon betont - für die Struktur der Gruppe, für
Untergruppenbildungen verantwortlich sind) sogar quantitativ diejenigen der Mutter
zu dem jeweiligen Individuum übertreffen. Der Sozialpartner der Mutter wird nun also
der Sozialpartner des Kindes. Bisher konnten wir entsprechendes nur zwischen
Individuen unterschiedlichen Geschlechtes beobachten, d. h. das junge Weibchen
präferiert den „Freund“ der Mutter, das junge Männchen hingegen deren
„Freundin“.
Durch Bubi wurde ich nicht nur zum vermeintlichen
„Affenzauberer50“,
vor allen ihm (und den anderen Kapuzinermännchen) verdanken wir auch die Klärung
einer Vokalisationsform, die im normalen Kapuzinerleben selten zu beobachten ist und
von uns als „Begrüßungsgeschrei“ bezeichnet (und von Nolte ([149]) richtig als
„Freudengeschrei“ interpretiert) wird. Sowohl Noltes Pablo als auch unser Bubi
verwenden es vor allem bei der Begrüßung ihnen vertrauter oder attraktiver
Personen.51
Es ist wohl derselbe Vokalisationstyp, den Dobroruka 1972 ([37]) als „the flute-like call
p-ru-pju-uiu-uiu-uiu-u“ beschrieben hat. Er wurde beobachtet als ein Kapuziner
entkam und nach der Annäherung an den Käfig (nach unserer Interpretation) seine
Artgenossen begrüßte. Weigel ([231]) schließlich ordnete den sogenannten „open mouth
staccato beep“ als Verteidigungsdrohen dem agonistischen Verhalten zu. Wir wissen
nun, welche Bedeutung dieser Vokalisationstyp hat. Normalerweise ist er zu
beobachten, wenn Männchen nach längerer physischer Trennung das erste Mal
wieder Kontakt bekommen, dann geben sie ihn (sehr laut) von sich, fassen
sich dabei eventuell auch gegenseitig an den Körper und an die Genitalien.
Auch Weibchen können sich an dieser Begrüßung beteiligen. Wir können ein
solches Geschrei experimentell hervorrufen, indem wir zwei Gruppen, die keinen
Gitterkontakt haben, Zugang zu benachbarten Käfigen verschaffen. In der
neuen Primatenstation (13.2.2) konnten wir es dann später täglich hören (s.
u.).
Interessante Einsichten in das Kapuzinerverhalten erbrachten auch ungewollte
Exkursionen unserer Tiere, die unermüdlich im Zerstören ihrer Käfiganlage relativ
häufig neue „Schwachstellen“ sichtbar machen, indem sie ihre Käfige verlassen und
den Haltungsraum erkunden, wobei sie nicht nur in der Lage sind, die Klimadecke zu
öffnen und für uns vorerst unsichtbar zu werden. Sie zeigen Versäumnisse der
Tierpfleger - eine nicht richtig verschlossene Tür oder aber auch das Vergessen nur
eines Sicherheitsschlosses - umgehend auf. Das Entkommen im Käfigraum führt zwar
zu erheblichen Zerstörungen, ist aber grundsätzlich unproblematisch, da es uns in der
Regel sehr schnell gelingt, den Flüchtling wieder einzufangen. Erwähnenswert ist hier,
dass einmal auch Hoppediz entkam und im Raum herumsprang. Ich war anwesend und
gab sofort die Anordnung, die noch offene Käfigtür zu schließen, um vorerst
alle anderen Affen zu sichern. Sekunden später sprang Hoppediz jedoch in
weiten Sätzen herbei, sprang über die Köpfe der vor dem Käfig Stehenden
an diesem herunter und öffnete seinen Käfig von außen und floh in diesen
zurück. Er hatte also offensichtlich nicht nur durch Versuch und Irrtum das
Öffnen des Käfigs gelernt, vielmehr hatte er das Prinzip „Türriegel“ völlig
verstanden.
Weitaus problematischer dagegen ist es, wenn sich die Außengehege als nicht gesichert
oder als nicht haltbar erweisen. Unsere ersten Kapuziner, die in das Freie entkamen,
drangen glücklicherweise in die Innenräume der Affenanlagen ein und besuchten
uns in der Küche. Auch die meisten anderen Exkursionen im Freien konnten
wir schnell beenden. Einmal war jedoch sechs Kapuzinern der Ausflug in die
Umgebung geglückt, so dass wir uns auf Kapuzinerfang einrichten mussten. Die
Kapuziner waren nämlich, zumindest teilweise, von dem Dach, auf dem sich die
Außengehege befanden, in die Umgebung entwichen. Glücklicherweise gelang
es uns, unterstützt von hilfreichen Mitarbeitern (wobei unser Hausmeister
Horst Schwedes sich besonders hervortat), drei Kapuziner sukzessive in Räume
der Hochschule zu jagen. Primus konnten wir so in einem Arbeitszimmer,
Jens im Eingangsbereich der Hochschule und Lisa in einem Fahrstuhl des
Mensagebäudes einfangen. Erwin wurde schließlich auch gesichtet, er untersuchte bzw.
zerstörte gerade eine ca. 30 Meter hohe Weide. Seine weiten Sprünge waren
wunderbar zu beobachten. Gleichzeitig wurde uns auch schon damals bewusst, wie
schwierig im Freiland das Beobachten sein dürfte, identifizierten wir doch den so
markanten unverwechselbaren Erwin nur sicher über eine Negativliste (alle
anderen adulten Männchen waren in der Anlage). Alle Versuche, Erwin zum
Absteigen zu bewegen, scheiterten. Schließlich mussten wir die Feuerwehr um
Hilfe bitten. Die Feuerwehrleute gingen davon aus, dass Erwin handzahm und
„herunterzuholen“ sei. Unser Anliegen, ihn „irgendwie“ zum Herunterklettern zu
bewegen, verstanden sie bald, überzeugte doch Erwin schnell, dass es nicht einfach ist,
einen adulten Kapuzinermann einzufangen, so sprang er in großen Sätzen von
einer Seite der Krone zur anderen. Schließlich gelang es den Feuerwehrleuten
durch ständiges gezieltes Anlegen der Leiter Erwin so zu bedrohen, dass er
den Abstieg bzw. den Absprung einem weiteren Verweilen vorzog. Sobald
Erwin den Boden erreicht hatte, gelang es uns, ihn in Richtung Hochschule zu
jagen, wo er in einem undurchdringlichen Gebüsch verschwand. Verstärkt
durch Kinder der Siedlung suchten wir nun das Gebüsch systematisch ab und
sichteten ihn auch mehrfach, obgleich er stets sehr schnell erneut unseren Blicken
entschwand. Schließlich war er so zornig und wohl auch erschöpft, dass er, statt zu
fliehen, uns angriff, wodurch einer der hilfsbereiten Feuerwehrleute ihn im
Netz fangen konnte. Der vierte Ausbrecher konnte zurückkehren. Die letzten
beiden noch fehlenden Kapuziner, Isabelle und Sarah, orteten wir schließlich in
unserem Kistenlager versteckt, es gelang uns, sie von dort sukzessive in das
Affenhaus zu jagen, wo wir sie schließlich in ihre gewohnten Räume zurückbringen
konnten.
Komplex gestalteten sich auch Lernexperimente. Studenten hatten die Aufgabe gestellt
bekommen, einfache Lernversuche durchzuführen. Zwei von ihnen, Cornelia Steiner und Elke
Erbe-Grünkorn52
dachten sich folgenden Versuch aus. Sie deckten zwei Futternäpfe mit Brettern ab, auf
denen eine bestimmte Anzahl von Punkten gezeichnet war. Gleichzeitig fertigten sie
Schilder, auf denen ebenfalls verschiedene Punkte angebracht waren. Ihre Idee
war folgende, sie wollten den Kapuzinern beibringen, dass die Punktzahl, die
sie hochhalten und zeigen, auch belohnt wird. Über die Komplexität des zu
protokollierenden Lernversuches waren sie sich nicht im Klaren, Einwände
fanden kein Gehör. Bereits in der ersten Sitzung griffen die Kapuziner wider
Erwarten signifikant richtig. Doch ab der zweiten Sitzung hatten sie gelernt,
dass es offensichtlich viel einfacher ist, beide Näpfe schnell hintereinander zu
öffnen, anstatt sich auf das Gezeigte zu konzentrieren. Cornelia Steiner und
Elke Erbe-Grünkorn benötigten dann allein ein ganzes Semester, um durch
mühevolle Änderungen am Versuchsaufbau, das Ergebnis des ersten Tages zu
verifizieren.
Wir schlossen unseren damaligen Befunde (1983) mit einem Ausblick: „In den
nächsten Jahren werden mit großer Wahrscheinlichkeit weitere Kinder, auch der
zweiten Generation in Menschenobhut, geboren und aufgezogen werden. Wir werden
also die Möglichkeit erhalten, zu prüfen, ob es so etwas wie Großeltern, Tante und
Onkel auch bei unseren Kapuzinern gibt bzw. ob diese für das Individuum
Bedeutung haben. Darüber hinaus werden Feinanalysen zeigen, wodurch die soziale
Stellung des Individuums bedingt wird und was für den Einzelnen attraktiv
und unattraktiv ist. In der nächsten Zukunft hoffen wir auch aufzeigen zu
können, welche Bedeutung gerade dem biologischen Vater zukommt und ob es
Paarbindungen unter Kapuzinern gibt oder nicht. Unabhängig von diesen
erhofften Resultaten der nächsten Zeit sammeln sich exakte lückenlose Daten
über Individualentwicklungen dieser hochinteressanten Affenart, die uns viele
Informationen über die Komplexität der Sozialstruktur liefern werden“ ([277],
Seite 125). Viele Antworten werde ich im folgenden Teil des Kapitels geben
können.
Bevor ich Daten aus der Primatenstation berichte, möchte ich noch Befunde zum
Vater bei Kapuzineraffen vorstellen, die wir in der provisorischen Haltung erhoben
hatten. Darüber hinaus gebe ich Informationen zu unserem Umgang bzw. zum
Umgang der Kapuzineraffen mit physischen Verletzungen.
Im August 1983 besuchte uns Leobert E. M. de Boer aus Rotterdam, gemeinsam mit Studenten
und erfahrenen Mitarbeitern des Rotterdamer Zoos. Gemeinsam mit mir fingen sie mit Ausnahme
von Kim53
alle Kapuzineraffen ein und entnahmen Blutproben für serogenetische Untersuchungen.
Diese Untersuchungen waren Bestandteil der Dissertationsarbeit von Frank
Princée ([169]). So hatten wir und speziell Petra Pippert hinreichendes
Datenmaterial für ihre Doktorarbeit ([153]) zur Bedeutung des „Biologischen
Vaters“ bei Kapuzineraffen. Für ihre Analysen standen ihr neben den Befunden
von Princée 12045 Viertelstundenprotokolle an Einzelindividuen und 3650
Halbstundenprotokolle an der Gesamtgruppe zur Verfügung. Zu diesem Zeitpunkt
bestand die Don-Gruppe aus 29, die Bubi-Erich-Hoppediz-Gruppen aus 17
Individuen. In unserer Hauptuntersuchungsgruppe, der Don-Gruppe, lebten
von den ursprünglich 17 Wildfängen (s. o.) noch zehn (4 Männchen, 6
Weibchen).54
In diesem Abschnitt folge ich den Befunden von Petra
Pippert.55
„Die Verwandtschaftsbeziehungen der in Kassel geborenen Jungtiere sind im Hinblick
auf Mutter und Geschwister (mütterlicherseits) bekannt. Wir gehen nämlich davon aus
- auch wenn wir die Geburt nicht beobachten konnten -, dass das Weibchen, das das
Neugeborene am Tag nach der Geburt - spätestens dann wird es entdeckt -
trägt und säugt auch die biologische Mutter ist. Da bisher pro Nacht nur
ein Junges geboren wurde, ist somit auch eine Verwechselung der Neonaten
ausgeschlossen.
Wir können jedoch kaum Aussagen über mögliche Väter,
Geschwister oder Halbgeschwister väterlicherseits wagen, da hierzu
Vaterschaftsausschlüsse56
geleistet werden müssten. Daher, insbesondere auch im Hinblick auf soziobiologische
Hypothesen (...), wo dem Investment des Vaters in die Nachkommen hohe
Bedeutung zugemessen wird, sollen im Folgenden mit Hilfe verschiedener Methoden
Vaterschaften ausgeschlossen bzw. wahrscheinlich gemacht werden. Hierzu
erscheint die Überprüfung der Solidität verschiedener Hypothesen der geeignete
Weg.
Gehen wir zunächst von der Hypothese aus, dass das Männchen, das zur
fraglichen Zeit mit der Mutter des zu untersuchenden Kindes kopuliert hat, bzw.
um das die Mutter zur fraglichen Zeit wirbt - also während des Oestrus, in
dem, bei Berücksichtigung der Tragzeitlänge, die Befruchtung stattgefunden
haben muss -, dass dieses Männchen der biologische Vater ist. Die sich hieraus
ergebenden Vaterschaften (beruhen) auf Kopulationen zur richtigen Zeit. (Petra
Pippert gibt hier eine Tabelle an, in der für 31 Kinder der so zu ermittelnde
Vater angegeben ist.) Nach diesen Daten wäre demnach das dominierende
Männchen (Bubi bzw. Don) das Tier, um das am meisten geworben wird,
bzw. das zur fraglichen Zeit mit dem Weibchen kopuliert. So müsste nahezu
ausschließlich dem jeweils dominierenden Männchen der verschiedenen Gruppen die
Vaterschaft zuerkannt werden. Dies verwundert nicht, ist doch nach unserem
Wissensstand das dominierende Männchen auch das attraktivste Tier der
Gruppe.


Wir beobachteten jedoch auch zweimal, dass Weibchen auch während eines Oestrus
mit verschiedenen Männchen kopulierten, bzw. um verschiedene Männchen warben
(...). Zudem können wir davon ausgehen, dass zwischen den Beobachtungen
Kopulationen stattfinden, die nicht protokolliert werden. Bei diesen sind die während
der regulären Befunderhebung nach Plan regelmäßig zu beobachtenden Interventionen
des dominierenden Männchens bei Kopulationen mit anderen Männchen nicht
zwingend zu erwarten, da den Tieren dann Innen- und Außengehege zur Verfügung
stehen (während der Beobachtungen befinden sich alle Tiere im Innenraum) und eine
Sichtbarriere besteht, die das dominierende Männchen daran hindert, alle
Geschehnisse in der Gruppe zu verfolgen. Diese Annahme wird durch zahlreiche
Zufallsbeobachtungen gestützt.
Daher wollen wir im Folgenden mit Hilfe moderner serogenetischer Methoden
überprüfen, ob eine Korrelation zwischen dem so ermittelten „Vater“ und dem
biologischen Vater besteht und wollen die entsprechenden Daten vergleichend
betrachten.
... Die Blutmuster wurden auf das Vorkommen elektrophoretischer Varianten
von 35 Enzymsystemen untersucht (vgl. [169]). Doch erwiesen sich für die
Vaterschaftsausschlüsse unserer Kapuzineraffen von den 35 Systemen nur
Glukosephosphatisomerase (GPI) und Mannosephosphatisomerase (MPI) als
verwendbar. (Petra Pippert gibt hier eine Tabelle an, in der für 24 Kinder die
möglichen Väter nach Princé ([169]) aufgelistet sind.)
Neben den so getätigten Ausschlüssen sind jedoch noch weitere Ausschlüsse möglich.
Bei dieser Tabelle ist ... noch nicht berücksichtigt, ob die entsprechenden Männchen
schon als Väter, ihrem Alter nach, in Frage kommen. Nach unseren eigenen Befunden
werden Kapuzinermännchen erst im Alter von 6-7 Jahren geschlechtsreif, ... Dass
Männchen in diesem Alter tatsächlich geschlechtsreif werden, belegt Odin, da hier
(nur) Erich oder Erwin als Väter in Frage kommen. Beide waren zum Zeitpunkt der
Zeugung 6-7 Jahre alt. ...
Eine weitere Reduktion (Eingrenzung) der möglichen Väter kann durch Berücksichtigung
der Gruppenhistorie, also von Zeugungstermin und Gruppenzusammensetzung
durchgeführt werden. ... Eine weitere Möglichkeit, Vaterschaften auszuschließen,
besteht darin, die Phänotypen von Kindern und Vätern zu vergleichen. ... Auf Grund
dieser phänotypischen Unterschiede ist Bubi eindeutig der biologische Vater der
Jungtiere Toko, Edith, Claudia, Florian und auch von Kim. ... Für die Individuen
Dido, Sebastian, Lea und Manuel müsste dementsprechend Bubi als möglicher Vater
entfallen (...), entsprechen sie doch voll dem Phänotyp von Cebus apella cay. ...
Insofern können wir nun den biologischen Vater für 8 Individuen sicher bestimmen
(...).
Die serogenetischen Befunde zeigen aber auch auf, wie problematisch es wäre, würden
wir dem (nach unseren sorgfältigen Beobachtungsprotokollen) jeweiligen Sexualpartner
der Mutter zur fraglichen Zeit (...) die Vaterschaft zuerkennen. Vergleichen wir
nämlich die (damals) neuen Daten mit den Ergebnissen der Untersuchungen zum
Verhaltensoestrus, dann stellen wir fest, dass die Angaben über den möglichen
biologischen Vater nicht immer übereinstimmen.
So kopulierte Sanga, die Mutter von Agnes und Lisa, mit dem adulten Männchen
Don, die Vaterschaft wird aber nach den Blutuntersuchungen den Männchen Iggo oder
Tschiggo zuerkannt. Auch Davids Mutter Dunka kopulierte zur fraglichen Zeit zwar
mit Don, von der Vaterschaft konnten aber alle Männchen außer Erwin ausgeschlossen
werden. Betrachten wir die Angaben für Odin, dann ist sein biologischer Vater Erich,
seine Mutter Migga warb jedoch im entsprechenden Oestrus um das dominierende
Männchen Bubi.
Insofern lässt sich für unsere Kapuzineraffen eindeutig falsifizieren, dass die
(beobachteten) Sexualpartner der Weibchen während der Oestren auch zwingend die
Väter der später geborenen Jungtiere sind. Demnach nutzen die Oestrusdaten
eigentlich noch nicht einmal, um bei mehreren möglichen Vätern einer Entscheidung
über die Vaterschaften näher zu kommen. Wir wollen daher nach anderen Wegen der
Vaterschaftseingrenzung suchen“ ([153], Seiten 21-30).
Petra Pippert ist dann der Hypothese nachgegangen, dass Väter verstärkt in ihre
Kinder investieren, und hat für jedes in Kassel geborene Individuum die über die Jahre
erhobenen Daten zur Sozialpartnerpräferenz sorgfältig analysiert. „Ziehen wir das
Resümee dieser Individualbefunde, so sind zwei Schlussfolgerungen schon jetzt
möglich: 1. Jedes Tier präferiert in jedem untersuchten Lebensjahr jeweils ein
Männchen. 2. Offensichtlich präferieren die einzelnen Jungtiere während der
(verschiedenen bis zu vier) untersuchten Lebensjahre jedoch kein bestimmtes
Männchen. Die Annahme, dass das jeweils präferierte Männchen auch der
Vater des entsprechenden Kindes ist, ist demnach falsifiziert“ ([153], Seiten
34-35).
„Für die Annahme des väterlichen Investments spricht daher ab dem dritten
Lebensjahr wenig. Damit ist jedoch noch keine Aussage über die ersten beiden
Lebensjahre gemacht. Hier müssen Gründe für das häufig auftretende eindeutige
Präferenzverhalten gesucht werden. Wir müssen zunächst einmal fragen, ob es der
Vater ist, der verstärkt mit seinen Kindern interagiert. (Petra Pippert konnte zeigen,)
... dass zumindest ein Drittel der nach dem Verhalten bzw. nach der soziobiologischen
Hypothese als Väter in Frage kommenden Männchen die biologischen Väter nicht sein
können. Bei den übrigen Jungtieren kommen zudem fast als Regel mehrere Männchen
als Väter in Frage, so dass die Annahme, der biologische Vater wäre zumindest bei den
restlichen zwei Dritteln auch Hauptsozialpartner, nicht hinreichend belegt werden
kann.
Wir müssen demnach nach anderen Erklärungen für die beobachteten Interaktionen
zwischen adulten Männchen und Kindern suchen. Überprüfen wir zunächst, inwieweit
das soziale Beziehungsgefüge der Mutter Einfluss auf die Interaktionspartner des
jeweiligen Jungtieres haben könnte. So unterhalten nämlich die adulten Weibchen,
unabhängig davon, um welches Männchen ein Weibchen während des Oestrus wirbt,
häufige Kontakte zu bestimmten adulten Männchen in der Gruppe. ... Vergleichen
wir nun die präferierten männlichen Sozialpartner der Mütter mit denen der
Kinder, dann stellen wir fest, dass sich diese Annahme - beide unterhalten zum
gleichen adulten Männchen bevorzugte Kontakte -, bei der Betrachtung der
quantitativen Befunde des ersten Lebensjahres bei 18 von 20 der für diese
Analyse zur Verfügung stehenden Jungtiere bewahrheitet (...)“ ([153], Seiten
39-40).57
Dem biologischen Vater kommt also bei den Kapuzineraffen keine Bedeutung
zu.58
Auch Kapuzineraffen können leider nicht lesen, sonst hätten sie sich vielleicht anders
verhalten.
Immer wieder habe ich von erheblichen Verletzungen berichtet, die sich bei Auseinandersetzungen im Sozialverband ergaben. Wie bereits in Kapitel 12 ausgeführt, durften verletzte Tiere im Sozialverband bleiben. Auf eine Behandlung habe ich bewusst verzichtet, hätte doch einer Behandlung mit verschiedenen Medikamenten umfangreiche Tierversuche an unverletzten Tieren vorausgesetzt, um die Risiken einer Applikation einschätzen zu können. Die Kapuzineraffen haben ein gutes Wundheilungsvermögen. Dieses unterstützen sie durch sorgfältiges Reinigen der Wunde. Hierzu setzten einige Tiere auch Werkzeug (Holzspäne) ein, um Eiter herauszupuhlen. In der Regel wurden die Wunden durch die Behandlung immer größer und flacher, dann heilte die Wunde vom äußeren Rand her sehr schnell zu. Ein „Wunder“ des Heilungsvermögens möchte ich nicht verschweigen: An einem Morgen im Jahr 1984 fanden wir Sanga mit fast völlig abgerissener linker Brust vor. Die Brust hing nur noch an einem Hautlappen. Wir haben Sanga sofort separiert und in einen kleineren Krallenaffenkäfig überführt - bei der Manipulationsbegeisterung der Kapuzineraffen war ein Verbleiben im Sozialverband nicht vertretbar gewesen, die anderen Gruppenmitglieder hätten ständig versucht, an der Brust zu ziehen. Nach der Separation rief ich sofort meine Tierärzte an und bat um Rat. Einmütig meinten diese, dass es zum Absetzen der Brust keine Alternative gebe. Überzeugt davon, dass dies Sanga besser kann als jeder Tierarzt, unternahm ich nichts. In den der Separation folgenden Wochen konnten wir fast täglich beobachten, wie die Brust sukzessive wieder anwuchs. Schließlich war Sanga „kosmetisch“ wieder völlig hergestellt. Ob die Milchsekretion noch richtig funktionierte, weiß ich nicht. Ich konnte aber Jahre später beobachten, dass ihr damals geborenes Jungtier an beiden Brustwarzen saugte und auch gesund aufwuchs. Natürlich gehen nicht alle Verletzungen gut aus, über einen dramatischen Verlust werde ich noch ausführlich berichten.
Meine Absicht war es, Kapuzineraffen sozusagen „von der Wiege bis zur
Bahre“ beobachtend zu begleiten. Anschließend wollte ich eine Monographie über
Gehaubte Kapuzineraffen schreiben. Dieses Forschungsprojekt zu beenden, ist
mir nicht vergönnt gewesen. Wir konnten aber über ein Jahrzehnt unsere
Forschungsergebnisse überprüfen und auf eine breitere Basis stellen. In der neuen
Primatenstation standen für die Kapuzineraffen sechs schon in den Kapiteln 11 und 12
beschriebene Haltungseinheiten zur Verfügung. Anfänglich hielten wir die Don-Gruppe
in drei dieser Einheiten (ohne trennende Gitter), die Hoppediz-Gruppe in
zwei Einheiten (mit trennendem Gitter), dazwischen eine Puffergruppe mit
Erich als dominierendem Männchen, um so den Gitterkontakt zwischen Bubi
und Don auszuschließen. Nach der Separation von Bubi (s. u.) gaben wir der
Don-Gruppe auch noch den vierten Käfig hinzu. In den Außengehegen standen
den Kapuzineraffen neben Sitzbrettern auch Klettergerüste zur Verfügung.
In den Innengehegen verzichteten wir auf zentrale Sitzgelegenheiten völlig,
da wir in der alten Haltung gelernt hatten, dass zentrale Strukturen ideal
sind, um andere Tiere zu bedrohen. Vielmehr brachten wir daher an den
Wänden übereinanderliegende, tiefe Ruhebretter an, so konnte ein bedrohtes
Individuum durch den Sprung auf das darunterliegende Brett leicht einem Angreifer
entkommen. Diese Anordnung war auch ideal für die Beobachter, konnten sie
doch von unserer Beobachtungsbank aus, alle Interaktionen im Sozialverband
protokollieren.
In dieser Zeit ist es uns gelungen, beide Gruppen zu erhalten. Wir haben verdrängte
Individuen entnommen und in anderen Räumen der Primatenstation gehalten. Zudem
habe ich später regelmäßig die jungen Nachzuchtmännchen im Alter von unter einem
Jahr an den Tierhandel abgegeben (um erkennbare Haltungsprobleme der Zukunft zu
vermeiden). Nur einmal habe ich ein Weibchen mit ihren Söhnen aus der
Don-Gruppe entfernt, da diese das relativ friedliche Zusammenleben massiv
gefährdeten.59
Die Kopfzahl beider Gruppen blieb über die Jahre stabil.
Grundsätzlich bestätigten die Kapuzineraffen unsere bisherigen Befunde mit
einer wichtigen Ausnahme. Auf Freundschaften zwischen Weibchen beruhende
Untergruppen gab es nicht mehr, wie an zwei Beispielen aufgezeigt sei. Teufel
war von 1978 bis 1982 vornehmlich mit Page befreundet, ab 1984 hielt sie
enge Kontakte nur noch zu eigenen Kindern. Sylvia war bis 1981 eng mit
Lena befreundet. Nach der Separation von Lena hielt sie enge Kontakte mit
der mit ihr importierten Dunka, danach nur noch mit ihren Kindern ([256]).
Untergruppenbeziehungen beruhten nun auschließlich auf Verwandtschaft
über die Mutter. Töchter blieben auf Dauer in der Untergruppe der Mutter,
Männchen bis zum Ende des fünften Lebensjahres. Danach reduzierten diese die
Kontakte zur mütterlichen Untergruppe nahezu vollständig und suchten sich
eine „Freundin“ aus dem Kreis der nichtvertrauten Individuen. Mit diesem
Weibchen hielten sie sehr engen Kontakt, paarähnliche Beziehungen waren die
Regel.
Darüber hinaus gab es in beiden Gruppen nicht mehr „den“ dominierenden Mann.
Man muss sich dabei vergegenwärtigen, dass unsere Sozialgruppen solche in
Menschenobhut waren. Unter Freilandbedingungen wäre es nie möglich gewesen, dass
ein Männchen - wie Don - über Jahrzehnte eine solche Position hält, vielmehr wäre es
schon mehrmals in dieser Zeit durch ein von Außen kommendes Männchen ersetzt
worden und hätte seine Gruppe zwangsläufig verlassen müssen. Selbst Don konnte
nicht sicher sein, der Vater der Kinder des dominierenden Weibchens zu sein. Sylvia
warb ab 1985 häufiger um Toko und kopulierte 1987 auch mit ihm während der
Beobachtungen ([256]).
Eine scheinbar berechtigte Kritik an der Forschung zur Bedeutung
verwandtschaftlicher Beziehungen in großen Sozialgruppen unter
Laborbedingungen und auch unter Freilandbedingungen in geschützten
Gebieten ist die Tatsache, dass unter „echten“ Freilandbedingungen die
Todesraten weit höher seien, dass es dort überhaupt kaum Verwandte geben
würde.60
Dieser Kritik möchte ich entgegenhalten, dass wir nicht wissen, wann in den
Millionenenjahren der Evolutionsgeschichte sich das Kümmern um Verwandte
(eigentlich um Vertraute ([277])) als selektionsbegünstigend herausgestellt hat. Dass
man dann, wenn es unter kargen Bedingungen kaum Verwandte (über die Mutter)
gibt, diese Präferenz nicht aufzeigen kann, ist banal. Ohne das Vorhandensein von
Kindern z. B. ist auch das Mutter-Kind-Verhalten nicht untersuchbar. Auch
dies gilt für viele Freilandgruppen, sterben doch manchmal alle Kinder. Hier
würde kein Kritiker widersprechen und behaupten, eigentlich gäbe es keine
Mutter-Kind-Beziehungen.
Beginnen möchte ich meinen Bericht mit Bubi. Bubi lebte nach einjähriger Trennung
seit 1983 wieder in der Hoppediz-Gruppe in einem stark gegliederten Gehege (s.o.). In
den Jahren der gemeinsamen Haltung kämpften Bubi und Hoppediz häufig
miteinander, vermieden aber den direkten physischen Kontakt. Sie kämpften durch ein
Gitter und vermieden offensichtlich die direkte Konfrontation. In der neuen
Primatenstation hielten wir die Hoppediz-Gruppe wiederum in einem durch ein Gitter
getrenntes Gehege. Diese Abtrennung hatte auch praktische Gründe. Bei widrigen
äußeren Bedingungen (Temperaturen unter -4 Grad) konnten wir die Kapuzineraffen
nicht aussperren. Durch dieses Gitter konnten wir sie dann „schieben“, die
Hoppediz-Gruppe in die linke Hälfte ihres Haltungskäfigs, die Erich-Gruppe in die
rechte Hälfte des Geheges und die Don-Gruppe in das Gehege der Erich-Gruppe.
Ein positives Nebenergebnis dieses Zwischengitters war die Vergrößerung der
Klettermöglichkeiten.61
Bubi nutzte dieses Zwischengitter wiederum zum Kämpfen mit Hoppediz. Beide saßen
sich - getrennt durch das Gitter - gegenüber und bedrohten sich gegenseitig massiv. Sie
wählten einen Platz an der Vorderfront, möglichst weit von der beide Seiten
verbindenden Schieberöffnung entfernt.
Die Situation in dieser Gruppe war nicht einfach, Hoppediz war eindeutig das
dominierende Männchen, doch hatten dies seine Weibchen (Migga, Inge, Purzel)
noch nicht erkannt. Besonders das dominierende Weibchen Migga schien zu
„glauben“, Bubi sei attraktiver als Hoppediz. Sie suchte die Nähe von Bubi
und putzte ihn. Seit Oktober 1983 (Tötung eines 26 Tage alten Jungtieres)
wuchs kein Jungtier in dieser Gruppe heran, auch die beiden 1985 in der
Gruppe geborenen Jungtiere wurden getötet. Wir haben das Töten nicht
beobachten können. Nach unserer Vermutung war Hoppediz der Töter. Da dieses
Männchen mit größter Wahrscheinlichkeit auch der Vater der Kinder war, Bubi
hatte kein Interesse am Geschlechtsverkehr, die Weibchen warben um Bubi
und kopulierten mit Hoppediz, hätte Hoppediz somit seine eigenen Kinder
getötet.62


Wäre die Hoppediz-Gruppe unsere einzige Kapuzineraffengruppe gewesen,
hätten wir sicherlich den vermeintlichen Übeltäter entfernt. Doch Hoppediz
durfte in der Gruppe bleiben. Die Weibchen putzten ihn dann 1985 auch
zunehmend, 1986 war er „der“ attraktive dominierende Mann, weitere Kinder
überlebten.
Meine Tierpflegerinnen hatten zunehmend Schwierigkeiten, Bubi zum Reinigen
auszusperren, ich ließ daher extra für Bubi noch eine Leiter einbauen, damit er auch
ohne Sprung den Durchgang nach Außen erklimmen konnte. Das Aussperren gestaltete
sich immer schwieriger. Am 28.12.1987 war es soweit, Bubi weigerte sich, nach außen
zu gehen, vielmehr blieb er auf dem Boden sitzen. Das Leben im Sozialverband war
nicht mehr zu verantworten.
Ich beschloss, Bubi in die Quarantäne umzusetzen, wollte ihn aber nicht einfangen.
Bubi musste von mir nur gerufen werden, er lief hinter mir her (durch den ihm nicht
vertrauten Flur und durch die Quarantäneschleuse) und ließ sich bewegen, einen der
dortigen Käfige zu betreten. Dort hielt er sich nur am Boden auf. Bubi hatte sich wohl
die Zukunft anders vorgestellt, mich würdigte er keines Blickes. Doch bereits am
29.12.1987 begrüßte er mich wieder mit lautem Geschrei. Jeweils am Morgen
und am Abend besuchten wir ihn bei unseren Rundgängen. Am 13.01.1988
erhielten wir dann unseren ersten Brüllaffen-Mann, der auch in die Quarantäne
untergebracht wurde (vgl. Kapitel 12). In den folgenden Tagen berichteten meine
Tierpflegerinnen, Bubi würde keine Nahrung mehr zu sich nehmen, wir müssten
wohl mit seinem Ableben rechnen. Was war geschehen? Wir alle waren von
unserem Brüllaffen begeistert, unser erster Gang war immer zu ihm. Bubi, im
Nachbarkäfig, war offensichtlich „beleidigt“. Nachdem ich dies erkannt hatte, änderten
wir unser Verhalten. Bei Betreten der Quarantäne begrüßten wir als erstes
Bubi und unterhielten uns mit ihm. Bubi genas und begrüßte mich bei dem
Eintreten wieder mit lautem Geschrei an der Käfigtür. Bubi blieb aber ansonsten
sehr bewegungseingeschränkt. Das an der Rückwand angebrachte Ruhebrett
konnte er offensichtlich nicht aufsuchen. Seine Haltung war ausgesprochen
unbefriedigend. Ich beschloss daher, ihn mit Weißbüschelaffen zu vergesellschaften. Das
Risiko für die Weißbüschelaffen schien mir vertretbar zu sein. Ich ließ zwei
Krallenaffenschlafkästen anbringen und überführte das am 25.03.1983 in Kassel
geborene Männchen C108 und seinen am 20.03.1987 geborenen Sohn C181 am
02.03.1988 in den Bubi-Käfig. Mir schien nur das echte Risiko zu bestehen, dass die
Weißbüschelaffen nicht genügend Vitamine bekommen würden. So ordnete
ich an, den Futternapf mit dem Vitaminbrei auf einen der Schlafkästen zu
stellen, unerreichbar für Bubi. Bei meinem Morgenrundgang bot sich mir ein
unerwartetes Bild. Der vermeintlich bewegungsunfähige Bubi sprang auf den
Schlafkästen offensichtlich „begeistert“ mehrfach hin und her, von einem
Schlafkasten auf den anderen, und hatte wahrscheinlich den Krallenaffenbrei
genossen.
Das Problem „Vitaminbrei“ war leicht zu lösen, ich ließ den Brei in einen der
Schlafkästen stellen. Bubi hielt sich in den folgenden Tagen nicht mehr am Boden auf,
er saß auf dem Ruhebrett, bei meinem Eintritt „flohen“ die Weißbüschelaffen zu
ihm und drängten sich an ihn. Es bestand also keine Notwendigkeit mehr,
Bubi in der Quarantäne abzusondern. Bereits am 07.03.1988 bezog Bubi ein
Krallenaffengehege (mit Zugang zu einem der Außengehege). Auch diesmal gingen wir
gemeinsam in das neue Gehege, Bubi folgte mir und konnte problemlos umgesetzt
werden.
Am 23.04.1988 tauschten wir ohne Probleme C181 durch das am 01.07.1985 geborene
Weibchen C140 aus. C108 und C140 erfreuten uns (und wohl auch Bubi) durch zwei
am 27.09.1988 geborene Jungtiere, die im Beisein des alten Kapuizinermannes
aufgezogen wurden. Die gemeinsame Haltung mussten wir erst aufgeben, nachdem wir
in diese gemischte Gruppe am 07.12.1988 Lea, die am 05.06.1981 geborenen
Inge-Tochter, introduzierten (Lea war aus der Hoppediz-Gruppe verdrängt
worden). Das erwachsene Kapuzinerweibchen vertrug sich zwar auch mit der
Weißbüschelaffenfamilie, doch passte es unerwartet auch in die Schlafkästen der
Krallenaffen und aß mit Begeisterung den Krallenaffenbrei auf. Daher gaben wir am
19.01.1989 die gemeinsame Haltung von Weißbüschelaffen und Kapuzineraffen auf.
Bubi lebte dann noch mehr als zwei Jahre gemeinsam mit Lea in der Primatenstation.
Bei seinem Tod am 03.03.1991 hatte ich nach sechzehn gemeinsamen Jahren einen
meiner besten Freunde verloren.
Wir schätzten alle Individuen unserer Sozialgruppen und litten mit ihnen. Besonders
belastete mich das unerwartete Schicksal von Sylvia. Daher möchte ich über dieses und
die Vorgeschichte berichten. Die Don-Gruppe lebte relativ friedlich zusammen. In der
Abbildung 13.79 habe ich das Beziehungsgeflecht in der Zeit vom 01.01.1992 bis
31.03.1993 skizziert. Die Skizze beruht auf 77652 protokollierten affiliativen Kontakten.
Diesen standen nur 272 agonistische Interaktionen gegenüber. Knapp 5 % der
affiliativen Kontakte (3047) waren Soziale Körperpflege. Da dieses Verhalten für
Präferenzverhalten der Individuen besonders informativ ist, gebe ich die präferierten
Partner jeweils an.
Das dominierende Männchen gab es nicht mehr. Es dominierte die Koalition von
Don, Christian und Toko, die in der Regel zentral gemeinsam zusammensaßen
und nach meiner Einschätzung bewusst jeglichen agonistischen Kontakt
vermieden.63
Alle drei Männchen saßen nicht nur gemeinsam zusammen, sie präferierten sich auch
untereinander bei der sozialen Körperpflege. Für Don waren Christian (n = 28) und
Toko (n = 9), für Toko Don (n = 56) und für Christian Don (n = 83) und Toko
(n = 15) die attraktivsten Sozialpartner bei dieser Verhaltensweise. Nur für
Toko sind Putzkontakte zu weiblichen Individuen bemerkenswert, er putzte
Sarah (n = 40), von der er 59 mal geputzt wurde. Sarah putzte auch Don (n
= 33) und Christian (n = 13), zudem sind die aktiven Putzkontakte von
Sarahs Töchtern Stephanie und Silke zu diesen Männchen bemerkenswert
(Stephanie: Don (n = 53), Toko (n = 50), Christian (n = 14); Silke: Don (n = 53),
Toko (n = 50), Christian (n = 14)), so dass Sarah nach dem protokollierten
Verhalten64
aus heutiger Sicht „das“ dominierende Weibchen war. Neben den Kontakten von
Sarah und ihren Töchtern zu diesen Männchen mag erwähnenswert sein, dass Sylvias
Tochter Sandra Christian dreizehnmal putzte.
Darüber hinaus mag man die große Untergruppe von Sylvia erkennen, die
als dominierende Untergruppe besondere Beziehungen zu diesen Männchen
unterhielt (s. o.). Die Matriarchin Sylvia unterhielt nur noch Beziehungen
gegenseitiger sozialer Körperpflege mit ihren Töchern Sarah (aktiv n = 21,
passiv n = 6) und Sandra (aktiv n = 18, passiv n = 19) und putzte auch ihre
Enkeltöchter Stephanie (n = 5) und Paula (n = 8). Sarah unterhielt neben den
Beziehungen zu ihrer Mutter gegenseitige Putzkontakte zu ihren Töchtern
Stephanie (aktiv n = 29, passiv n = 12), Silke (aktiv n = 25, passiv n = 35) und
Franziska (aktiv n = 36, passiv n = 12), zudem putzte sie ihren Sohn Sophiles (n
= 14), ihre Enkeltochter Tonia (n = 26) und ihre Nichte Paula (n = 29).
Sarahs Tochter Stephanie hatte neben den Kontakten mit ihrer Mutter Beziehungen
gegenseitiger sozialer Körperpflege mit ihrer Schwester Silke (aktiv n = 17, passiv n = 43),
ihrer Tochter Tonia (aktiv n = 57, passiv n = 34) und putzte auch die Petra-Tochter Paula
(n = 14).65
Daneben erkennt man vier weitere Untergruppen, nämlich (1) Teufel und ihre Kinder und
Enkelkinder66,
(2) Kim mit Kindern und „Freunden“ (Kim gelang es, neben den Beziehungen
gegenseitiger sozialer Körperpflege zu ihren Töchtern Karin (aktiv n = 14, passiv n =
53) und Karoline (aktiv n = 23, passiv n = 12) auch solche mit den nicht über die
Mutter verwandten adulten Männchen Carlos (Mutter: Erna) (aktiv n = 30, passiv n
= 32) und Justus (aktiv n = 22, passiv n = 37) zu etablieren; Karin war
nach den Putzkontakten nur mit Carlos befreundet (aktiv n = 67, passiv n
= 16), putzte aber auch ihre Schwester Karoline (n = 12)), (3) Edith mit
Kindern67 und (4)
Claudia mit Kindern68.
Untergruppenübergreifende Putzkontakte konnten zu Teufel festgestellt
werden (sie wurde von Kim (n = 11) und Claudia (n = 4) geputzt,
zudem putzte Claudia Edith (n = 7). Mit quantitativ vernachlässigbaren
Sozialkontakten lebten in der Gruppe ein weiterer Sohn Ernas, Cesar, und
Dorothee69.
Dorothee wurde von Karoline (n = 8) geputzt und putzte selber den peripher lebenden
Philipp (n = 8)
Ich spekulierte damals, dass irgendwann die Gruppe nur noch aus dem Sylvia-Clan
bestehen würde, war dieser doch besonders mächtig. Das dominierende Weibchen
Sylvia schien unangreifbar, ihr standen ihre Töchter Sarah und Petra verläßlich zur
Seite. Ich sollte mich täuschen. Irgendwie erlitt Paula, die am 20.05.1991 geborene
Tochter von Petra, eine Verletzung, an der die sie beständig pflegende kerngesunde
Petra für uns völlig unerwartet am 01.02.1992 starb. Nach dem Autopsiebericht hatte
sie sich bei der Wundreinigung mit Eitererregern infiziert. Die Pflege von Paula
übernahmen dann die weiblichen Verwandten.
In der Gruppe wurden zwischen Mai und Juli fünf weitere Jungtiere geboren und
aufgezogen.70
Am 02.11.1992 wurde Sylvias Sohn Samuel massiv von Justus angegriffen und verletzt
und starb am selben Tag. Sylvia hatte also nach Petra einen zweiten möglichen
Verbündeten verloren. Am 08.01.1993 schließlich griff plötzlich Stephanie
Sylvia massiv an. Sylvia hatte keine Hilfe. Sarah half nicht gegen die eigene
Tochter, sonstige Freunde hatte Sylvia - als (ehemals) dominierendes Weibchen -
nicht, sie war den Attacken ihrer Enkeltochter hilflos ausgesetzt. Offensichtlich
war die Mutter-Tochter-Bindung für Sarahs Handeln entscheidender als die
Tochter-Mutter-Bindung. Stephanie kämpfte unerbittlich. Ich habe dann die wehr- und
schutzlose Sylvia aus der Gruppe genommen und wegen ihrer schweren physischen und
psychischen Verletzungen eingeschläfert.
Toko, der erste in Kassel gezeugte Kapuzineraffe, war ein Ausnahmetalent.
Er übernahm oder erbte (was ich vermute) das Verhalten seiner Mutter
Teufel, gegen sich gerichtete Drohungen zu ignorieren und stattdessen
in die gleiche Richtung wie der Drohende zu drohen, er beobachtete auch
sehr sorgfältig unser Verhalten. In der Primatenstation hatten wir einen
großen Beobachtungsraum, von dem aus wir auch außerhalb der regulären
Beobachtungszeiten, das Geschehen in der Don-Gruppe innen und außen begleiten
konnten.71
Doch auch die Kapuzineraffen konnten uns beobachten, wie wir gemeinsam
Kaffe tranken und auch feierten. Besonders aufmerksam war hier Toko, sobald
ich aufstand, um den Kapuzineraffen etwas abzugeben, erkannte er dies und
wartete dann bereits am Gitter auf die Gabe. Toko kooperierte auch bei
Versuchen von Elisabetta Visalberghi zum Werkzeuggebrauch ([227]) und
„las ihr jeden Wunsch von den Augen ab“, er klopfte jede Nuß, die er
mühelos mit den Zähnen hätte öffnen können, mit einem der angebotenen
Bretter.72
Zudem war er auch ein verlässlicher Partner bei allen Bedrohungen, ob Ratte oder
Weihnachtsmann. Toko zeigte niemals ängstliches Verhalten. Nach dem in Kapitel 11
beschriebenen Unfall rief mich Elisabetta Visalberghi an und fragte, war es Toko, was
ich leider nur bejahen konnte.
Zur bereits mehrfach erwähnten Edith, meiner „Tochter“, möchte ich noch berichten, dass sie in den vielen Jahren als einziger Kapuzineraffe niemals vor mir gewichen ist, alle anderen respektierten mich und wichen vor mir, betrat ich das Gehege. Ich hatte seit ihrem Rücktransport in die Tierhaltung niemals den Versuch gemacht, Edith zu berühren, bin aber sicher, dass sie sich sehr wohl an die Aufzuchtzeit erinnert hat, dass sie wohl auch durch diese Phase in der Zeit, in der ein normal aufwachsender Kapuzineraffe die Zahl seiner Sozialpartner sukzessive ausweitet (s. u.), auf den Menschen geprägt worden ist. Ich hatte im Gehege der Don-Gruppe (mit Edith im Rücken) ein gewisses Gefühl der Sorge, sie könnte mich unkontrolliert angreifen. Hier war sie aber in den vielen gemeinsamen Jahren ein verlässlicher Partner.

Neugeborene Kapuzineraffen73 sind völlig abhängig von der Mutter und werden vornehmlich von dieser auf dem Rücken getragen, im ersten Lebensmonat quer zur Körperlängsachse, später längs zur Körperlängsachse.
Nur selten ist das Tragen am Bauch zu beobachten. Das Tragen durch andere
Gruppenmitglieder kann nur selten beobachtet werden. Nur eines der Kinder (Cesar,
vgl. Tabelle 13.3) wurde regelmäßig von seiner damals vierjährigen Schwester Claudia
(vgl. Tabelle 13.3) getragen. Von den ingesamt beobachteteten 547 Fällen des Tragens
durch Nichtmütter entfallen auf Claudia 325 Trageepisoden. Grundsätzlich waren die
Haupttragtiere Geschwister, vor allem Schwestern (73 %). Andere Juvenile
transportierten die Kinder ebenfalls (männliche und weibliche zu jeweils 7 %. Das
Tragen durch Adulte war selten zu beobachten (adulte Weibchen 1 %, adulte
Männchen 2 %).
Ab dem zweiten Lebensmonat werden die Kinder zunehmend alleine angetroffen, doch
bleiben sie bis zum Ende des vierten Lebensmonates für mehr als die Hälfte der
Beobachtungszeit auf oder neben der Mutter. Bereits in dieser frühen Lebensphase
sind Geschlechtsunterschiede aufzeigbar, männliche Kinder verlassen die Mutter früher
als weibliche Kinder.
Bereits während des ersten Lebensmonates beginnen die Kinder auf der Mutter
herumzuklettern, ab dem zweiten Lebensmonat richten sie sich auch auf, lösen also den
Kontakt mit den Händen. Ab dem dritten Lebensmonat klettern, laufen und springen
sie allein, wobei das Klettern (am Käfiggitter) am häufigsten zu beobachten ist. Auch
bei diesen frühen vorsichtigen Bewegungen sind Geschlechtsunterschiede aufzeigbar,
männliche Kinder springen ab dem vierten Lebensmonat signifikant häufiger als ihre
Altersgenossinnen.
Während der ersten sechs Lebensmonate werden die kleinen Kapuzineraffen immer
attraktiver für andere Gruppenmitglieder. Auch die Kinder beginnen, mit diesen aktiv
zu interagieren. Schematisch betrachtet (Abbildung 13.96), können wir feststellen, dass
die ersten zwei Lebensmonate - die jungen Affen sind noch völlig hilflos -
charakterisiert sind durch passive Kontakte mit Geschwistern, wobei ältere Brüder
erst ab dem zweiten Lebensmonat an dem Geschwister interessiert sind. Im
dritten Lebensmonat suchen dann die Kinder selbst den Kontakt zu ihren
Geschwistern. Im vierten Lebensmonat werden sie attraktiv für ältere im gleichen Jahr
geborene Jungtiere, die sie dann im fünften Lebensmonat auch selber aktiv
aufsuchen.74
In diesem Lebensmonat beginnen auch adulte Männchen, Kontakt zu den Kindern
aufzunehmen. In der Regel ist dies aber nur eines der adulten Männchen, häufig das
dominierende Männchen, das dann auch im sechsten Lebensmonat von den Kindern
aufgesucht wird. Dieses Männchen hatten wir dann den psychischen Vater genannt (s.
o.). Im sechsten Lebensmonat weiten die Kapuzinerkinder auch erstmals den Kreis der
Sozialpartner aktiv aus, sie nehmen auch Kontakte auf zu nichtverwandten ein Jahr
älteren Jungtieren.
Gerade im Hinblick auf unsere ersten Beobachtungen an importierten
Kapuzineraffenkindern ist es interessant, die Häufigkeit bestimmter Verhaltensweisen
im ersten halben Lebensjahr zu betrachten, wir werteten hierzu die Verhaltensweisen
Gehenzu, Manipulieren, Zusammensitzen ohne Körperkontakt, Kontaktsitzen, Soziales
Spiel, Soziale Körperflege aus und zusätzlich das Lippenschmatzen und das Schmusen
aus. Bei der letztgenannten Verhaltensweise betasten sich die Sozialpartner
gegenseitig und haben dabei einen „verliebten“ Gesichtsausdruck, der dem
charakteristischen Oestrusgesicht der erwachsenen Kapuzineraffen gleicht. Bei dem
Zusammensitzen (mit und ohne Körperkontakt), dem Sozialen Spiel und dem
Schmusen konnte zwischen aktivem und passivem Partner nicht differenziert
werden.

Bei dem aktiven Gehenzu (im Mittel 441 ± 247) zeigen die Kinder kein klares
Präferenzverhalten. Sie bevorzugen jedoch adulte Männchen, Geschwister und im
gleichen Jahr geborene oder ein Jahr ältere Juvenile und gehen seltener zu adulten und
juvenilen (nichtverwandten) Weibchen. Bei dem passiven Gehenzu (797 ± 562) sind sie
besonders attraktiv für Geschwister. Zudem ist auffällig, dass Kinder des gleichen
Altersbereiches (gleichalt und 1 Jahr älter) andere Kinder häufiger aufsuchen als
andere Gruppenmitglieder.
Geschwister und Kinder des gleichen Altersbereiches sind auch die bevorzugten Partner
bei dem Zusammensitzen mit (328 ± 178) und ohne (218 ± 178) Körperkontakt,
bei dem Sozialen Spiel (363 ± 178) und dem Schmusen (81 ± 43). Bei dem
Zusammensitzen ohne Körperkontakt sind zudem die Kontakte zu adulten Männchen
auffällig.75
Bei dem Manipulieren (aktiv: 48 ± 27, passiv: 116 ± 66) sind die Kinder eindeutig die
passiveren Partner, dies gilt noch weit auffälliger für die Verhaltensweise Soziale
Körperpflege (aktiv: 1 ± 1, passiv: 221 ± 102) und Lippenschmatzen (aktiv: 5 ± 4,
passiv: 172 ± 59). Hier sind es nahezu ausschließlich Geschwister und ein Jahr ältere
Juvenile, die aktiv mit den Kindern interagieren.
Geschlechtsunterschiede sind in dieser Altersphase gering. Nur bei dem sozialen Spiel
konnten wir diese sichern, männliche Kinder spielen signifikant häufiger als weibliche
Altersgenossen.76
Das zweite halbe Lebensjahr ist von besonderem Interesse, da die jungen Affen
zumindest theoretisch den Kreis der bevorzugten Sozialpartner nach ihrer eigenen
Wahl erweitern können. In dieser Lebensphase wird der kleine Kapuzineraffe
fortwährend immer unabhängiger von der Mutter und interagiert häufig mit anderen
Gruppenmitgliedern. Er wird attraktiv als passiver und/oder aktiver Partner. Das
einzige Verhalten, das die Kapuzineraffen auch im zweiten Lebenshalbjahr nicht
zeigen, ist die Soziale Körperpflege, doch sind sie häufig passive Putzpartner. Bei
unseren Analysen vergleichen wir die gemittelten Ergebnisse jeweils mit dem
Erwartungswert, mit dem Wert, der zu erwarten wäre, wenn das jeweilige Kind mit
jedem Indivduum seiner Gruppe gleich häufig interagieren würde. Im Vergleich zu den
Befunden des ersten halben Lebensjahres geben wir keine Daten zum Lippenschmatzen
an, da dieses Verhalten im zweiten Lebenshalbjahr nur sehr selten zu beobachten ist.
Zudem verzichten wir auf Daten des Zusammensitzens ohne Körperkontakt, da in
dieser Altersphase und unter den restriktiven Bedingungen in Menschenobhut nicht
entschieden werden kann, ob bestimmte Individuen zufällig oder absichtlich
zusammensitzen.77
Auch während des zweiten halben Lebensjahres bleibt die Mutter der wichtigste
Sozialpartner ihres Kindes. Dies gilt besonders bezüglich der Verhaltensweisen
Soziale Körperpflege (das Kind ist nur passiver Partner), Manipulieren und
Kontaktsitzen. Insofern ist besonders auffällig, dass die Kinder mit ihren
Müttern seltener spielen als zu erwarten. Geschlechtsunterschiede bei den
Mutter-Kind-Interaktionen sind nicht aufzeigbar. Doch sind weibliche Kinder in
dieser Lebensphase als passive Partner bei der sozialen Körperpflege und
bei dem Manipulieren für die Mutter attraktiver als für irgendein anderes
Gruppenmitglied.78
Bei dem Gehenzu79
ist auffällig, dass männliche und weibliche Kinder im zweiten Lebenshalbjahr
bestimmte andere Individuen präferieren, nämlich adulte Männchen, Geschwister und
Kinder des gleichen Altersbereiches. Männliche Kinder sind weniger daran interessiert
adulte und juvenile Weibchen und mehr daran interessiert subadulte und - schwach
gesichert - juvenile Männchen aufzusuchen im Vergleich zu weiblichen Kindern. Als
passive Partner werden männliche Kinder häufiger von Geschwistern und im gleichen
Jahr geborenen Kindern und seltener von adulten und juvenilen Weibchen aufgesucht
als weibliche Kinder.
Bei dem Manipulieren80
waren Kinder beiderlei Geschlechts als aktive und als passive Partner für Kinder ihres
Alterbereichs attraktiver als zu erwarten. Männliche Kinder waren zudem relativ
attraktiver für subadulte und juvenile Männchen. Diese Befunde waren aber nur
schwach zu sichern.
Bei dem Kontaktsitzen81
sitzen die Kinder weitaus häufiger mit Geschwistern und im gleichen Jahr geborenen
Kindern in Körperkontakt als zu erwarten. Weibliche Kinder sind zudem als
Kontaktpartner attraktiv für ein Jahr ältere Juvenile. Vergleichen wir die
Daten männlicher und weiblicher Kinder, gewinnt man den Eindruck, dass
männliche Kinder attraktiver für subadulte und juvenile Männchen und weniger
attraktiv für adulte Männchen sind als weibliche. Von diesen Ergebnissen
kann die hohe Attraktivität der Geschwister für männliche und weibliche
Kinder, von im gleichen Jahr Geborenen für männliche Kinder - verglichen mit
den Erwartungswerten - und die höhere Attraktivität adulter Männchen für
weibliche Kinder - verglichen mit männlichen Kindern - statistisch gesichert
werden.
Neben den Geschwistern spielen Kinder desselben Altersbereichs und - in geringerem
Maße - juvenile Männchen mit den 6 bis 12 Monate alten Kindern häufiger als zu
erwarten. Dies kann für beide Geschlechter für im gleichen Jahr geborene Kinder, für
männliche Kinder bezüglich der Geschwister und für weibliche Kinder bezüglich der ein
Jahr Älteren gesichert werden. Zusätzlich und bezogen auf alle Sozialpartner spielen
männliche Kinder signifikant häufiger als weibliche Kinder. Wir protokollierten das
soziale Spiel 1000 ± 158 mal für männliche und 383 ± 97 mal für weibliche Kinder im
zweiten Lebenshalbjahr.
Bei dem Schmusen82
sind Kinder desselben Altersbereichs die bevorzugten Partner. Geschwister beteiligen
sich häufiger als zu erwarten.
Im auffälligen Kontrast zu den bisher besprochenen Verhaltenweisen sind die Kinder bei der Sozialen
Körperpflege83
vor allem passive Partner. Darüber hinaus ist auffällig, dass es eigentlich nur die
Geschwister sind, die - unabhängig vom Geschlecht des Kindes - die Kinder
putzen. Weibliche Kinder sind hier auch attraktiv für adulte und juvenile
Weibchen.
Fassen wir die bisherigen Befunde zusammen, dann sind im zweiten Lebenshalbjahr
neben der Mutter nur drei Kategorien von Sozialpartnern für die Kapuzineraffenkinder
von Bedeutung, nämlich Geschwister, im gleichen Jahr geborene Jungtiere und ein
Jahr ältere Individuen. Dabei kommt auch dem Geschlecht der Sozialpartner hohe
Bedeutung zu. Bei den Kontakten zu ein Jahr älteren Jungtieren bevorzugen
männliche Individuen männliche und weibliche weibliche. Besonders auffällig ist dies
bei weiblichen Jungtieren. Ein Jahr ältere Männchen bevorzugen bei dem sozialen
Spiel, dem Manpulieren und dem Schmusen eindeutig männliche Kinder. Männliche
Kinder bevorzugen ein Jahr ältere Männchen bei dem Gehenzu, weibliche Kinder ein
Jahr ältere Weibchen bei dem Gehenzu, dem Kontaktsitzen, dem Schmusen und
umgekehrt.
Die gegenseitige Bevorzugung von Individuen des eigenen Geschlechtes ist besonders
auffällig bei dem Sozialen Spiel, Männchen spielen bevorzugt mit Männchen und
Weibchen mit Weibchen.
Die einzige Ausnahme von der Bevorzugung von Individuen des eigenen Geschlechtes
zeigen ältere Schwestern bei dem Gehenzu, Kontaktsitzen und bei der Sozialen
Körperpflege, hier präferieren sie männliche Kinder, was sich aber statistisch nicht
sichern läßt. Ältere Brüder hingegen interagieren mit den Kindern unabhängig von
deren Geschlecht (Kontaktsitzen) oder bevorzugen männliche Kinder (Soziales Spiel,
Manipulieren, Soziale Körperpflege und Schmusen), wobei sich diese Bevorzugung nur
für das Schmusen sichern lässt.
Zudem muss bemerkt werden, dass ältere Schwestern generell mit den Kindern
häufiger in engem Körperkontakt sitzen als ältere Brüder. Dies gilt auch für das
Manipulieren und besonders für die soziale Köperpflege, wo Schwestern die einzigen
erwähnenswerten sozialen Partner der Kinder sind. Dies gilt für männliche und
weibliche Kinder.
Betrachten wir nur die Beziehungen der im gleichen Jahr geborenen Kinder
untereinander, dann schmusen weibliche Kinder bevorzugt mit weiblichen Kindern,
manipuliert werden dagegen sowohl von weiblichen als auch von männlichen Kindern
bevorzugt gleichalte weibliche Artgenossen.
Auffällig ist insgesamt betrachtet, dass die sich im ersten Lebenshalbjahr
sukzessive entwickelten Sozialkontakte zu anderen Gruppenmitgliedern nicht weiter
ausweiten.
Zu Beginn unserer Untersuchungen war es offen, inwieweit die engen Mutter-Kind-Beziehungen
beibehalten werden würden bzw. ob auch im späteren Leben Unterschiede zwischen
Mutter-Sohn- bzw. Mutter-Tochter-Beziehung aufzeigbar sind. Die guten Zuchterfolge in
Kassel und die hohen Überlebensraten erlaubten quantitatives Datenmaterial auswerten zu
können.84
Wir verglichen auch hier die Befunde mit dem Erwartungswert. Neben den
prozentualen Anteilen analysierten wir die absoluten Häufigkeiten des zu
beobachtenden Verhaltens.
Männliche und weibliche Juvenile suchen ihre Mutter signifikant häufiger auf als zu
erwarten. Töchter sind bei dem Gehenzu signifikant aktiver als Söhne. Auch im 5.
Lebensjahr ist dieser Trend zu belegen. Diese geschlechtstypischen Unterschiede gelten,
unabhängig davon, ob man die prozentualen Anteile oder die absolute Häufigkeit des
aktiven Verhaltens Gehenzu betrachtet.
Söhne und Töchter werden von ihren Müttern durchgängig häufiger aufgesucht als zu
erwarten. Ab dem vierten Lebensjahr werden Töchter häufiger aufgesucht als Söhne,
was sich nach den absoluten Werten aber erst im fünften Lebensjahr sichern läßt, d. h.
Geschlechtsunterschiede sind bei dem Aufgesuchtwerden durch die Mutter sehr
gering.
Das am häufigsten zwischen den Müttern und ihren heranwachsenden Kinder zu
beobachtende Verhalten ist das Kontaktsitzen. Unabhängig vom Geschlecht und Alter
des Kindes sitzen die Mütter in engem Körperkontakt mit diesen.
Ab dem zweiten Lebensjahr putzen die juvenilen Kapuzineraffen ihre Mütter häufiger
als andere Gruppenmitglieder, unabhängig vom Alter und Geschlecht der Juvenilen.
Töchter putzen ihre Mütter häufiger als Söhne, was sich ab dem dritten Lebensjahr
sichern läßt. Sowohl bei den Töchtern als auch bei den Söhnen nimmt die absolute
Häufigkeit des Putzens der Mutter mit zunehmendem Alter zu (bei den Söhnen auf
einem geringeren Niveau).
Die Mutter wiederum putzt ihre Kinder unabhängig vom Geschlecht und
Alter signifikant häufiger als andere Gruppenmitglieder. Männliche Juvenile
sind für die Mütter dabei relativ attraktiver als weibliche. Das Interesse der
Mutter, die Jungen zu putzen, nimmt mit zunehmendem Alter der Juvenilen ab.
Darüber hinaus ist auffällig, dass Töchter bei der Sozialen Körperpflege in
der Mutter-Tochter-Dyade mit zunehmendem Alter die aktiveren Partner
werden.
Als Partner des Sozialen Spielens sind Mütter für ihre heranwachsenden
Kinder ohne Bedeutung, mit Söhnen spielen sie sogar signifikant seltener als zu
erwarten.
Für Juvenile Kapuzineraffen bleibt die Mutter der wichtigste Sozialpartner.
Verwandtschaftsbeziehungen (über die Mutter) sind also von hoher Bedeutung
für das im Sozialverband heranwachsende Individuum. Mit zunehmendem
Alter der Juvenilen werden Unterschiede zwischen der Mutter-Sohn- und der
Mutter-Tochter-Dyade immer auffälliger. In der Mutter-Sohn-Dyade bleibt die Mutter
der aktivere Partner, in der Mutter-Tochter-Dyade hingegen wird die Tochter die
aktivere. Hierdurch scheinen Töchter die engen Beziehungen zu ihren Müttern
sicherzustellen und zu erhalten, wodurch Matriclane und auf Verwandtschaft
beruhende Untergruppen in der Kapuzinergesellschaft geformt werden. Männchen
hingegen reduzieren mit zunehmendem Alter ihre Kontakte zur mütterlichen
Untergruppe (und suchen dann Kontakte außerhalb des Kreises der mütterlichen
Verwandten).
Bei den Beziehungen der Geschwister untereinander wollten wir untersuchen, ob diese
Beziehungen stabil bleiben oder sukzessive zurückgehen, zudem ob die Beziehungen von
Geschwistern des gleichen Geschlechtes sich von Bruder-Schwester-Dyaden unterscheiden
lassen.85
Ohne hier näher auf diese Befunde eingehen zu wollen, war durchgängiges und gut
gesichertes Ergebnis bei dem aktiven und passiven Gehenzu und bei dem
Kontaktsitzen, dass Geschwister über die gesamten vier betrachteten Lebensjahre ihre
Geschwister - unabhängig vom Geschlecht- häufiger aufsuchten bzw. von diesen
aufgesucht wurden und häufiger mit diesen in Körperkontakt saßen als zu
erwarten.
Bei der Sozialen Körperpflege muss einschränkend bemerkt werden, dass Brüder
grundsätzlich nur selten andere Individuen putzen bzw. dass Schwestern erst ab dem
dritten Lebenjahr häufiger aktiv andere Sozialpartner putzen. Männchen und
Weibchen werden von ihren Geschwistern, vor allem von ihren Schwestern, in allen
untersuchten Altersstufen signifikant häufiger geputzt als von irgendeinem anderen
Gruppenmitglied. Dagegen werden Weibchen nur selten von ihren Brüdern geputzt,
stets seltener als von ihren Schwestern.
Bei dem Sozialen Spiel spielen männliche Juvenile signifikant häufiger mit ihren
Brüdern (nicht aber mit ihren Schwestern) als zu erwarten. Weibliche Juvenile spielen
wenig.
Sie spielen jedoch mit Geschwistern unabhängig vom Geschlecht häufiger als mit
anderen Gruppenmitgliedern (Schwester/Schwester: 2. Lj. p < 0,05, 3. Lj. p < 0,005, 4.
Lj. p < 0,001, 5. Lj. p < 0,005; Schwester/Bruder: 4. Lj. p < 0,025, 5. Lj. p < 0,05).
Auch bei weiblichen Juvenilen ist eine Präferenz für weibliche Spielpartner
aufzeigbar, insofern spielen alle Juvenilen bevorzugt mit Individuen des eigenen
Geschlechtes.
Unabhängig vom Alter interagieren in der Gruppe geborene juvenile Kapuzineraffen
mit ihren (über die Mutter verwandten) Geschwistern, unabhängig auch von dem Grad
der Verwandtschaft (über den Vater). Sie suchen sich gegenseitig auf und sitzen
häufiger in engem Körperkontakt. Entsprechendes gilt für die soziale Körperpflege und
das soziale Spiel, doch bevorzugen sie hier gleichgeschlechtliche Geschwister.

Hella Kröger [115] untersuchte zudem die Kontaktsitzdyaden von zehn
Kindern der zweiten Generation. Neben den drei noch in der alten Station
geborenen Kindern Edgar, Cris und Stephanie (vgl. Tabelle 13.3) analysierte
sie die Daten von Silke (geb. 18.06.1986, Mutter: Sarah), Clarence (geb.
28.07.1986, Mutter: Claudia), Jan (geb. 02.07.1987, Mutter: Jonaine), Vic (geb.
23.07.1987, Mutter: Isabelle), Sophiles (geb. 31.08.1987, Mutter: Sarah), Ralf
(geb. 29.10.1987, Mutter: Edith) und Pia (geb. 14.11.1987, Mutter: Petra).
„Ich86
beschränke mich hier auf Daten zum Kontaktsitzen, da diese Verhaltensweise nach den
bisherigen Befunden besonders informativ ist. Da zwei Kinder (Edgar und Ralf) von einer
handaufgezogenen Mutter geboren wurden, berichte ich vorerst nur die Befunde von acht der
zehn Kinder.87
Erwartungsgemäß unterhalten auch die Kinder der zweiten Generation signifikant
häufiger enge Kontakte zur Mutter als zu erwarten, was für beide Geschlechter belegt
werden kann.
Neben denjenigen zur Mutter sind die Beziehungen zu Geschwistern wiederum
besonders eng, alle Kinder saßen mit ihren Geschwistern häufiger in Kontakt als zu
erwarten. Wegen der geringen Anzahl der möglichen Dyaden (drei) kann dieser
Unterschied nicht für beide Geschlechter getrennt voneinander belegt werden,
wenngleich jedes dieser Kinder mit seinem Geschwister viel häufiger interagierte als zu
erwarten.
Darüber hinaus unterhielten die Kinder engeren Kontakt zu ihrer Großmutter, ihren
Tanten - unabhängig vom Alter der Tanten - und ihren juvenilen (nicht älter als fünf
Jahre) Onkeln. Diese Unterschiede können auch separat für beide Geschlechter
gesichert werden, wobei bei den Beziehungen zwischen Nichten und juvenilen Tanten
wegen der kleinen Anzahl der möglichen Dyaden (zwei) statistische Tests nicht möglich
sind, auch wenn beide untersuchten Weibchen mit ihren juvenilen Tanten häufiger als
zu erwarten interagierten. Zudem sollte bemerkt werden, dass Verwandte aller vier
Qualitäten (Großmutter, adulte Tante, juvenile Tante, juveniler Onkel) mit
männlichen verwandten Kindern häufiger in engem Körperkontakt saßen als mit
weiblichen, was nach den bisherigen Befunden eigentlich auch zu erwarten
ist.
Subadulte und adulte Onkel waren jedoch als Kontaktpartner für die Kinder der
zweiten Generation ohne Bedeutung. Dies gilt besonders für die einzige zu
analysierende Nichte-subadulter Onkel-Dyade.
Die Befunde über Beziehungen mit Cousins und Cousinen beruhen nur auf wenigen
Individuen, wobei in keinem Fall das jeweilige Kind seltener mit einem Cousin oder
einer Cousine in engem Körperkontakt saß als zu erwarten.
Somit sind für männliche und weibliche Kinder des gehaubten Kapuzineraffen die
weiteren Verwandten ebenfalls wichtige Sozialpartner. Durch diese Beziehungen wird
das Kind Mitglied des mütterlichen Matriclans. Da - wie bereits betont - männliche
Nachkommen mit zunehmendem Alter die Kontakte zu ihrer mütterlichen
Verwandtschaft reduzieren, verwundert es nicht, dass ältere Männchen für die Kinder
der zweiten Generation keine Bedeutung als Sozialpartner haben“ ([115], Seiten
110-115).
Bereits mehrfach habe ich über meine „Tochter“ Edith berichtet, sie war das optimale
Versuchstier zum Verwandtenerkennen, lebte sie doch zwei Jahrzehnte in einer
Sozialgruppe mit ihrer biologischen Mutter und ihren Geschwistern. Bei der
Vergesellschaftung mit den Individuen ihrer Sozialgruppe hatte sie anfänglich -
wie berichtet - überhaupt keine Sozialkontakte. Dies verwundert nach den
vorgestellten Befunden nicht mehr. Im Alter von einem halben Jahr - dem Alter von
Ediths Introduktion - haben Kapuzineraffenkinder die Sozialisation bereits
abgeschlossen. Edith musste sich also auf Spielkontakte beschränken und
spielte mit ihrem Bruder Toko und den väterlichen Halbschwestern Kim und
Claudia.88
Zur Wertung dieser Befunde wollte ich auf verlässliche Befunde der Langzeitstudie
warten. Die entsprechenden Befunde hat dann Hella Kröger ([115]) ausgewertet.
„Edith konnte mit Individuen interagieren, die ihr alle gleich wenig vertraut waren.
Während ihres bisherigen Lebens interagierte Edith durchgängig häufiger mit
nichtverwandten als mit genetisch verwandten Individuen, insbesonders wird dies
deutlich, wenn wir ihre Kontakte zur biologischen Mutter mit denjenigen
einer ihrer Schwestern vergleichen, die von ihrer Mutter selbst aufgezogen
wurden. ... Betrachten wir nun die Beziehungen von Ediths Kindern mit anderen
Gruppenmitgliedern, dann ist bemerkenswert, dass die Großmutter, die Tanten und die
juvenilen Onkel als Sozialpartner ohne Bedeutung sind. Subadulte und adulte
Onkel dagegen interagieren mit ihren Neffen häufiger als zu erwarten. Insofern
machen diese Befunde deutlich, dass es nicht die genetische Verwandtschaft ist,
die für die engen Beziehungen zwischen Verwandten verantwortlich ist, es
ist vielmehr die Vertrautheit, die wichtig für die Beziehungen zwischen den
Gruppenmitgliedern einer wachsenden Sozialgruppe ist. In einer normalen
Sozialgruppe sind sicherlich nahe Verwandte auch vertraut miteinander, wobei
Kapuzineraffen
aber offensichtlich nicht in der Lage sind, verwandtschaftliche Beziehungen ohne Vertrautheit zu erkennen. Diese Annahme wird zudem durch die engen Beziehungen der zwei männlichen Kinder mit ihren subadulten und adulten Onkeln gestützt. Unter normalen Umständen würden diese nicht mit ihren Neffen interagieren, da ihre Schwestern als Sexual- und Kontaktpartner uninteressant sind. Die handaufgezogene Mutter, ein nachgewiesenes Vollgeschwister eines der Männchen und ein mütterliches Halbgeschwister des anderen, war dagegen als Kontakt- und Sexualpartner für beide Männchen attraktiv. Durch diese Kontakte interagierten sie auch mit den Kindern dieses Weibchens, unabhängig vom Grad der genetischen Verwandtschaft“ ([115], Seiten 121-124).
„Im Rahmen einer Längsschnittstudie konnte exaktes Datenmaterial durch
Einzeltierbeobachtungen (jeweils stand nur ein Individuum im Focus des Beobachters)
an in den Sozialgruppen aufwachsenden Jungtieren gesammelt werden. Als Beispiel
seien hier die Ergebnisse von zwei dreijährigen Kapuzineraffen, von Daniel
und Claudia, vorgestellt. Beide lebten in derselben Sozialgruppe gemeinsam
mit ihrer Mutter und jeweils einem damals einjährigen Bruder. Betrachten
wir ihre gesamten sozialen Interaktionen während des vierten Lebensjahres,
ist auffällig, dass jeder Kontakte zu verschiedenen Individuen der Gruppe
unterhält.
Bei den Mutter-Kind Interaktionen ist auffällig, dass Claudia ihre Mutter (13) vor
allen anderen Sozialpartnern präferiert: Daniel interagiert mit seiner Mutter (19)
weitaus seltener, wenngleich häufiger als mit irgendeinem weiteren adulten Weibchen
der Gruppe (16, 09, 12, 13, 18, 02). Besonders sei darauf verwiesen, dass Claudia
in der Mutter-Kind-Dyade die aktivere ist, Daniel hingegen der passivere
Partner. Beide, Claudia und Daniel, interagieren häufig mit ihrem jeweiligen
Bruder (38 bzw. 37), Claudia zudem noch mit juvenilen Weibchen. Besonders
bemerkenswert sind hier ihre Beziehungen zu zwei ein Jahr älteren Weibchen (22, 23).
Darüber hinaus unterhält sie Kontakt zu einem der adulten Männchen (07),
einem „Freund“ ihrer Mutter. Daniel dagegen interagiert vornehmlich mit
juvenilen Männchen, meist mit zwei jüngeren (31, 38) und einem älteren (21)
Männchen.
Betrachten wir die Qualitäten der Interaktionen, so gibt uns das bloße
Zusammensitzen (braun) nahezu keine Information. Bei dem Zusammensitzen in
engem Körperkontakt (grün) wird deutlich, dass Claudia dieses Verhalten weitaus
häufiger als Daniel zeigt, bei dem Sozialen Spiel (gelb), dass Daniel häufig
und Claudia selten spielt. Das Verhalten Gehenzu (blau) gibt keine neuen
Informationen, wenngleich auffällig ist, dass beide nahezu genauso häufig von
den Partnern aufgesucht werden, zu denen sie selbst den Kontakt suchen.
Weitere geschlechtstypische Unterschiede werden jedoch bezüglich des Verhaltens
Soziale Körperpflege (rot) auffällig. Daniel unterhält Putzkontakte nur mit der
eigenen Mutter. Bemerkenswert sind zudem die Unterschiede zwischen beiden
Individuen bei den Beziehungen zum eigenen Bruder, Daniel spielt mit seinem
Bruder, Claudia hingegen sitzt in engem Körperkontakt mit diesem und putzt
ihn.
Im vierten Lebensjahr ist also bereits das geschlechtstypische Verhalten ausgeprägt.
Die Wahl der Sozialpartner und die Qualität der Interaktionen erlauben, Männchen
und Weibchen voneinander zu diskriminieren“ ([255], Seiten 26-29).
„Ein unbeabsichtigter Kontrollversuch erlaubt, den Einfluss der Beobachter auf die
protokollierte Häufigkeit bestimmter Verhaltensweisen zu testen. Das Geschlecht
eines in der Gruppe geborenen Tieres wurde falsch bestimmt und das kleine
Kapuzineraffenmädchen (35) als Männchen „Jonas“ (vgl. Tabelle 13.3) jahrelang
beobachtet.89
Erst eine Gravidität im sechsten Lebensjahr (Jonas wurde zu Jonaine) erlaubte, den
Irrtum zu korrigieren.
Zwar war dieser Kapuzineraffe kein „Focus-Tier“, Protokolle von Einzeltierbeobachtungen
lagen nicht vor, doch wurde neben den Beobachtungen an bestimmten Individuen der
Gruppe auch die gesamte Gruppe täglich eine Stunde lang protokolliert. Das
vorliegende Datenmaterial erlaubt also zu fragen, ob das vermeintliche Geschlecht
Einfluss auf das Protokollieren hat. Zum Vergleich werden Daten eines im gleichen
Jahr geborenen Männchens, Carlos, angegeben. (Wir vergleichen also die Daten von
zwei Individuen, die über vier Jahre als Männchen parallel beobachtet worden sind.
Betrachten wir vorerst nur das
Kontaktsitzen90,
so ist bemerkenswert, ..., dass Carlos dieses Verhalten in den ersten drei Lebensjahren
häufiger zeigt als Jonaine, im vierten Jahr jedoch sitzt Jonaine häufiger mit ihrer
Mutter im Körperkontakt.
Bei dem sozialen Spiel jedoch sind vom ersten Lebensjahr an die zu erwartenden
Geschlechtsdifferenzen auffällig. Carlos spielt weitaus häufiger als Jonaine, beide
entsprechen diesbezüglich dem bisherigen Wissensstand.
Betrachten wir nun die Befunde zur sozialen Körperpflege und im speziellen die
Putzkontakte mit der eigenen Mutter, dann ist auffällig, dass beide im ersten
Lebensjahr ausschließlich passive Partner sind. Während Carlos durchgehend
der passive Partner bleibt, putzt Jonaine ab dem dritten Lebensjahr ihre
Mutter häufiger als sie von dieser geputzt wird. Wir erhalten das gleiche
Ergebnis wie bei den Einzeltierbeobachtungen und können somit den Einfluss
der Beobachter auf das Ergebnis abschätzen, ein unbeabsichtigtes selektives
Beobachten kann offensichtlich ausgeschlossen werden“ ([255], Seiten 32-33).
Nach der Separation von Bubi entfiel die Notwendigkeit, Gitterkontakt zur
Don-Gruppe zu vermeiden, wir konnten dieser Gruppe (unserer wichtigsten
Sozialgruppe) eine weitere Haltungseinheit zur Verfügung stellen. Die zwei kleinen
Gruppen (Hoppediz-Gruppe und Erich-Gruppe) hielten wir parallel. Sie dienten hier
1987 und 1988 Martina Pec als Datenlieferanten für ihre Dissertation ([152]). Martina
Pec berichtet in ihrer Arbeit, dass Erich für seine Weibchen (Erna und Lena) nicht
attraktiv war, diese warben durch das Gitter um Don. Daher beschlossen wir, die
beiden kleinen Gruppen zu fusionieren, was gelang. Unter dem Druck der
mächtigen Nachbargruppe waren auch Hoppediz und Erich gemeinsam zu
halten.
Die beiden Kapuzineraffengruppen hatten sowohl in den Innen- als auch in den
Außengehegen ständigen Gitterkontakt, was sicherlich eine wichtige Voraussetzung
für das relativ friedliche Zusammenleben in beiden Gruppen war und was
nach meiner Einschätzung die gemeinsame Haltung so vieler erwachsener
Männchen ermöglichte. Die Männchen beider Gruppen kämpften regelmäßig
gemeinsam gegen die „Feinden“ in der Nachbargruppe. Bei den Beobachtungen
konnte man den Eindruck gewinnen, das Zusammentreffen beider Gruppen
mit direktem Kontakt würde zu sofortigen massiven Beschädigungskämpfen
führen. Während der jahrelangen Haltung geschah es gelegentlich, dass das
Sicherheitsschloss, das den Verbindungsschieber sicherte, den ständigen
Rüttelversuchen91
der Kapuzineraffen nicht standhielt, so dass die Männchen beider Gruppen die
tatsächliche Möglichkeit der Konfrontation hatten. In solchen Fällen stoppte der laute
Kampf völlig, die großen kampfbereiten Männchen zogen sich sofort - lautlos - in die
entferntesten Bereiche ihres Geheges zurück, so dass wir sehr leicht ein neues Schloss
anbringen konnten.
Alle Affen in der Primatenstation wurden am Abend in die Innengehege gesperrt und erhielten
erst am Morgen Zugang zu den Außengehegen bzw. wurden zum Reinigen der Innengehege
ausgesperrt92
und durften erst nach dem Abtrocknen die Innengehege wieder betreten. Diese
Routine nutzten die Kapuzineraffen beider Gruppen zum lauten Begrüßungsgeschrei,
zweimal am Tag. Sie begrüßten sich am Morgen im Außengehege und am späten
Vormittag im Innengehege. Sowohl das laute Kämpfen während des Tages als auch
das laute Begrüßungsgeschrei dürften für die Kapuzineraffen wichtig gewesen
sein.
In den Tabellen 13.4 und 13.5 habe ich die Zusammensetzung der beiden
Kapuzineraffengruppen Ende 1996 angegeben.
In der Hoppediz-Gruppe (Tabelle 13.4), in der aber Hoppediz nicht mehr dominierte,
Florian und Manuel waren attraktiver für die Weibchen dieses Sozialverbandes ([39]),
lebten noch vier der ursprünglich importierten Tiere. Insgesamt bestand die
Gruppe im November 1996 aus 17 Individuen, sechs adulten Männchen, vier
adulten Weibchen, 5 juvenilen Weibchen und zwei Kindern. Sowohl Erna als
auch Purzel waren noch reproduktiv aktiv. Purzel hatte eine sehr periphere
Stellung.93
In der Hoppediz-Gruppe war Migga das dominierende Weibchen. 1995 erkämpfte sich
Erna (unterstützt durch ihre Tochter Kristina) diese Position, Migga und Dido
mussten wir separieren. Ihre erwachsenen Söhne waren keine Hilfe. Die nach diesen
Konflikten geborene Erna-Tochter Elka wurde von Eusebia am Tag nach der Geburt
adoptiert und aufgezogen ([39]). Katharina Klewitz beobachtete die Hoppediz-Gruppe
in den letzten zwölf Wochen vor der Entfernung aus der Primatenstation
(Mitte April 1997). Purzels Position hatte sich relativ verbessert, da nun auch
Diana94
ohne weibliche Verwandte sich mit einer Randposition abfinden musste. Katharina
Klewitz verglich sorgfältig die soziale Entwicklung der beiden in der Gruppe lebenden
Kinder.95
Ihre Hypothese, Purzels Stellung würde sich wieder mit zunehmender Selbständigkeit
verschlechtern, konnte nicht mehr überprüft werden. Jedenfalls bestand die
Hoppediz-Gruppe vor der Abgabe im Wesentlichen aus den Weibchen des Erna-Clans
und den adulten Migga-Söhnen, wobei bei den Augsburger Kapuzineraffen (s. o.)
phänotypisch Migga besonders repräsentiert war.
Sowohl an der Hoppediz- als auch an der Don-Gruppe erhob Michael Dulitz auch
umfangreiches Datenmaterial zu Änderungen des Verhaltens im Verhaltensoestrus und
nach Geburten ([39]), die an die bisherigen Befunde anschließen.
In der Don-Gruppe (Tabelle 13.5) lebten Ende 1996 36 Individuen, mit Ausnahme von Don und Teufel waren alle Gruppenmitglieder bei uns geboren. Dreizehn männlichen Gruppenmitgliedern, acht von ihnen adult, drei Kinder. Von den 23 weiblichen Individuen waren 13 adult. Fragen wir nach der Repräsentanz unserer Gründerweibchen, dann lebten weibliche Nachkommen von Sylvia, Teufel, Ama und Erna in diesem Sozialverband. Dabei ist eigentlich der relativ hohe Anteil von Amas Nachkommen überraschend, hatte doch Ama selber nur eine Tochter (Kim) hinterlassen. Don und Teufel waren sehr alt, dennoch aber äußert mächtige Gruppenmitglieder. Wie bereits ausgeführt (s. o.) hatten die Clan-Mütter Kim und Claudia auch über Teufel Sozialkontakte. Wie alte Menschen sahen Teufel und Don jedoch nicht mehr „jung und knusprig“ aus, ihr Haarkleid war schütter geworden. Insofern wollten die von Verantwortlichen der Hochschule ausgesuchten neuen Halter diese beiden Persönlichkeiten nicht übernehmen. Die Verantwortlichen gaben meine beiden wunderbaren Freunde verantwortungslos als Beschäftigungsobjekte für demente Alte in einem Altenheim ab. Unabhängig davon, dass beide nicht zahm, vielmehr ausgesprochen wehrhaft waren, sind Kapuzineraffen als Spielzeuge ungeeignet. Beide hätten ein besseres Schicksal verdient.
In den Medien tauchen immer wieder angeblich zahme Kapuzineraffen auf, als
„helfende Hände“ wird ihnen fast der Status von Hilfspersonal zuerkannt. Diese
Kapuzineraffen sind aber hart trainierte Individuen, die mit hohem Aufwand
begleitet werden müssen. Sie haben tatsächlich auch eine wichtige Funktion
für Hilfsbedürftigte mit schweren Gebrechen. Diese werden durch die Affen
ansprechbarer, wir Mitmenschen können mit dem Hilfsbedürftigen über den Affen
kommunizieren, wir können ihn nach seinem Helfer befragen, wodurch Frager und
Befragter profitieren. Man kann sich darüber sicherlich streiten, ob dieser
Umstand hinreichend ist, um den hohen finanziellen Aufwand für entsprechende
Forschungsprojekte zu rechtfertigen. Ohne Zweifel tut man dem trainierten Affen
keinen Gefallen.
Auf dem Weg zum Verstehen einer Kapuzinergesellschaft bin ich in den Jahren in
Kassel weit vorangekommen, gesicherte Angaben zur Menopause und zum Höchstalter
konnte ich nicht mehr leisten. Unsere Individuen mit bekannter Vorgeschichte sind
leider zu beliebigen Kapuzineraffen verkommen.

1Der Text des ersten Teils des Berichts über Kapuzineraffen ist streckenweise wortidentisch mit einer 1979 veröffentlichten Arbeit ([241]) und einem Beitrag, den ich gemeinsam mit Petra Pippert und Cornelia Witt in der Zeitschrift des Kölner Zoo 1983 veröffentlicht habe ([277]).
2Angela Nolte stellte mir auch ihre Bilddokumente ihrer Kapuzineraffen Pablo, Suse und Bambino zur Verfügung. Einige dieser Bilder werden Bestandteile meines Berichtes sein.
3Für ihr Entgegenkommen danke ich Frau Direktor Dr. Barbara Jantschke.
4Die besten Beobachtungsgebiete gibt es nach Aussagen von Kollegen in der Nähe von nur mit dem Hubschrauber erreichbaren Ölförderungsfeldern.
5Zwei der Individuen wurden in Kassel in der Hoppediz-Gruppe geboren, Erna trug bei uns den Namen Kristina, Purzel den Namen Plato.
6Auch bei Kapuizineraffen kann man sich bei der Geschlechtsbestimmung täuschen (vgl. Abb. 13.39).
7Die Entwicklung des Sozialgefüges heranwachsender Kapuzineraffen, Cebus apella cay, unter besonderer Berücksichtigung des ersten halben Lebensjahres. Staatsexamensarbeit (1983)
8Zu den sozialen Beziehungen männlicher und juveniler Kapuzineraffen, Cebus apella, unter der besonderen Berücksichtigung von Verhaltensänderungen bei Oestren und Geburten. Staatsexamensarbeit (1984).
9Brinkmann, C.: Zum Sozialverhalten von Cebus apella cay Illiger, 1815 in Gefangenschaft: Die experimentelle Bildung einer Gruppe. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe an der Geamthochschule Kassel. Kassel, 15. September 1978.
10Pippert, P.: Zur Entwicklung des sozialen Beziehungsgefüges heranwachsender Kapuzineraffen Cebus apella cay (ILLIGER, 1815) in Gefangenschaft. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe und die Oberstufe an der Gesamthochschule Kassel. Kassel, 28. April 1982.
11Schulz, S.: Soziale Beziehungen adulter Kapuzineraffenweibchen, Cebus apella, unter besonderer Berücksichtigung des Einflusses der Östren und Geburten auf die Stabilität des Sozialgefüges. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe an der Gesamthochschule Kassel. Kassel, 20. September 1983.
12Dulitz, M.: Parameter des sozialen Beziehungsgefüges in einer etablierten Sozialgruppe des Gehaubten Kapuzineraffen Cebus apella. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe und die Oberstufe an der Universität Gesamthochschule Kassel. Kassel, 11. April 1995.
13Klewitz, K.: Der Einfluss der Sozialpartnerwahl der Mutter auf die Sozialpartnerwahl des Kindes in einer etablierten Sozialgruppe des Gehaubten Kapuzineraffen Cebus apella. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt an Gymnasien. Kassel, 20. Februar 1998.
14Leider muss ich bekennen, dass das Filmen eine problematische Form der Befunderhebung ist. Die Auswertung der Filmaufnahmen dauert extrem lange, zudem „sieht“ man selbst bei unserer aufwendigen Befunderhebung mit zwei Kameras gleichzeitig (Übersichtsbild und geführte Kamera mit Zoom) weit weniger als ein trainierter Beobachter. Dies waren die Gründe, warum ich später in der Primatenstation trotz optimaler Bedingungen (Videoanschluss in jedem Raum, optimierte Führung der Kameras) dann völlig auf Videoaufzeichnungen verzichtet habe.
15Meinel war bewusst, dass man Primaten nur dann verantwortbar verbrauchen könne, wenn man diese selber züchten würde.
16Die Tatsache, dass es sich bei den Tieren um Babies handelte, war uns damals aber noch nicht bewusst. Heute bin ich sicher, hätten wir uns über die berechtigten restriktiven Importbestimmungen hinweggesetzt und die Babies einzeln und mit engem Kontakt zum Menschen „gehändelt“, hätten mehrere Individuen eine Überlebenschance gehabt. Freilich wäre dies mit Gesundheitsgefahren für den menschlichen Betreuer verbunden gewesen. Diese Risiken habe ich persönlich nie gescheut. Bei zahlreichen Importen (vor dem Neubau der Primatenstation) fand die Quarantäne dann in meinem Privathaus statt, so wurden die Mitarbeiter in der Tierpflege (und die übrigen Primaten) vor möglichen Infektionen bewahrt.
17Meine Hauptaufgabe in dieser Zeit war es, die neuen Räumlichkeiten überhaupt „benutzbar“ zu machen. Die Räume wurden (trotz aufwendiger Klimaanlage) nicht warm, hier schaffte ein Umbau der Abluftkanäle Abhilfe, wir verlegten diese auf den Boden, so dass die warme Luft nun von der Klimadecke durch den jeweiligen Haltungsraum „hindurchgesogen“ wurde.
18Der Käfig 5 war unterteilt.
19Diese Forderung war nicht risikolos, sollte es doch nicht möglich sein, erwachsene neu importierte Kapuzineraffen am Leben zu halten: „Alte Kapuziner wollen sich nicht an die Gefangenschaft gewöhnen: sie werden traurig, verschmähen, Nahrung zu sich zu nehmen, lassen sich niemals zähmen und sterben gewöhnlich nach wenigen Wochen ([68], Seite 490).
20Bei dem zweiten Import fielen die geschwächten Importtiere auf den gefliesten Käfigboden, ich konnte nicht ausschließen, dass auch der Absturz zum Tod der Tiere beigetragen hatte.
21Nach neuester Systematik [141] wären heute unsere Cebus apella cay „richtig“ als Sapajus cay zu bezeichnen, Bubi wäre einer anderen Art, Sapajus libidinosus, zuzuordnen. Unsere Zuchterfolge der Vergangenheit, über die ich berichte, erlauben berechtigte Zweifel, ob diese systematische Aufspaltung auf Artenniveau berechtigt ist.
22Die Individuen dieses Transportes werden im Text als B-Tiere bezeichnet.
23Bei der Fusion beider Gruppen am 19. August 1975 war auffällig, dass gleich nach Öffnen des
Verbindungsschiebers alle Tiere der A-Gruppe die Außenkäfige aufsuchten, während die Männchen
der B-Gruppe und die Weibchen Erna und Ama den Käfig der A-Gruppe (Käfig V der
Haltungsskizze) untersuchten. Erste soziale Kontakte waren nur zwischen den Männchen
beider Gruppen zu beobachten; sie beschränkten sich auf Aufreiten und Kampfspiele,
anfangs waren hier die Männchen der B-Gruppe aktiv, 15 Minuten nach Fusion stieg jedoch
auch Tschiggo bei Iggo auf. Am Abend des folgenden Tages waren sechs Schlafgruppen zu
beobachten, nämlich (1) Bubi/Hoppediz, (2) Tschiggo/Purzel, (3) Teufel/Inge/Primus, (4)
Baja/Migga, (5) Erich/Iggo/Erna, (6) Erwin/Sanga/Ama, die über alle fünf Innenkäfige verteilt
waren.
In den folgenden Tagen bildeten sich drei Untergruppen, nämlich (1) eine reine Männchenuntergruppe,
bestehend aus Bubi/Tschiggo/Hoppediz, also allen Männchen der A-Gruppe, und den drei
neu hinzugekommenen Männchen Erich/Erwin/Iggo - wobei bemerkenswert ist, dass kein
B-Gruppen-Männchen bzw. kein B-Gruppentier zu Bubi aktiv sozialen Kontakt aufnahm bzw.
aufnehmen konnte, (2) eine reine Weibchenuntergruppe Ama/Sanga/Migga, wohl auf Sanga zentriert,
zu der auch das solitäre Weibchen Baja Kontakt aufzunehmen versuchte; (3) Teufel/Inge/Primus;
daneben die beiden Weibchen Purzel und Erna, die relativ unattraktiv für andere Gruppenmitglieder
waren und Tendenzen zur Männchenuntergruppe zeigten).
Zwischen den Individuen einer Untergruppe konnten keine agonistischen Aktivitäten beobachtet
werden. Agonistische Interaktionen richteten sich besonders gegen die Nur-Weibchengruppe.
24Bei den parallel gehaltenen und beobachteten Javanermakaken (Kapitel 14) dagegen wirkte jede Kopulation wie eine Vergewaltigung, insofern sympathisierten meine Mitarbeiterinnen mit den Kapuzineraffen.
25„In this species cycling females show active courtship behavior; they closely follow the males searching for proximity and body contact. During courtship, female’s facial expression consists of raised eyebrows and forehead, ears drawn back flat against the head and mouth corner drawn backward. Furthermore, she vocalizes and looks for eye-to-eye contact with the partner. Her most important aim, however, is to make the male follow her. In absence of this response no copulation will occur. Especially at the beginning, the male can be reluctant. Female courtship can last hours and even days without being reciprocated, and can also fail. In general, however, the female’s persistent and continuous attempts to elicit her partner’s interest are successful and a coordination of their behavior patterns is attained. Eye-contacts, play (cuff, chase and approach-withdrawal interactions), arm raised and chest caressing sitting in front of each other, body contact, become more and more frequent prior to copulation“ ([228], Seite 164).
26Diese wollte aber mit Makaken arbeiten und war an den Kapuzineraffen - im Nachhinein glücklicherweise - nicht interessiert.
27Als zweifelhaftes Geschenk erhielt ich also zusätzlich ein kleines kräftiges und gesundes Kapuzinermädchen, das bei uns in Grifte aufgezogen wurde. Über Edith und den durch dieses Weibchen ermöglichten Erkenntnisgewinn werde ich noch mehrfach berichten.
28Wir hatten statt drei nun fünf Kapuzineraffengruppen.
29In der damaligen Kapuzineraffenhaltung hatten wir mehrere nach unten offene Schlafkästen installiert, in denen sich bedrängte Tiere zurückziehen konnten, diese haben sich aber nicht bewährt, vielmehr konnten diese zur Falle werden.
30Zu gefährlichen Auseinandersetzungen kommt es nur, wenn sich in einem Gehege zwei Männchen begegnen, die sich für dominierend halten. Dann entscheidet ein Kampf, wer endgültig dominiert. Das unterlegene Männchen gibt jedoch nicht auf. Im Freiland dürfte es auswandern, in Menschenobhut müssen wir es - um Verluste zu vermeiden - separieren.
31Diese Entscheidung musste ich alleine treffen, meine Mitarbeiter waren gegen diese Introduktion, sie wollten Bubi separat halten.
32Gerade Bubi war bis zur Ankunft von Don unbestritten dominierend. Er blieb es für die übrigen Kapuziner: Ein physisch kräftiges, unbestritten dominierendes, jungadultes Männchen (Iggo) lebt gemeinsam mit seiner Gruppe seit Monaten in einem Gehege. Plötzlich betritt ein altes, fast bewegungsunfähiges, zudem verletztes Männchen den Käfig. Der Ausgang dieses Experimentes scheint klar. In der Tat aber kletterte Bubi mühselig auf ein hochgelegenes Brett, während Iggo auf den Boden sprang und stereotyp im Kreis herumzulaufen begann. Er hielt sich nicht für dominierend und gab sofort auf. Aus der Iggo-Gruppe wurde die neue Bubi-Gruppe.
33Die Gruppen bezeichneten wir immer nach dem jeweils dominierenden Männchen.
34Besonders intensiv jagten und verfolgten Teufel Sylvia und Primus Jens, zu dem Primus bisher keinerlei Kontakt aufgenommen hatte. In dem Haltungsraum drohten darüber hinaus auch alle Tiere der Erwin-Gruppe den neu importierten Kapuzinern. Nach dem Einsperren der Don-Gruppe in die Innenkäfige stellten sich sofort die alten Konstellationen wieder ein. Zwei weitere Versuche erbrachten das gleiche Ergebnis: kollektives Drohen fast aller Tiere der A- und der B-Gruppe gegenüber Don, wobei nur Bubi sich nicht beteiligte, Verfolgen von Sylvia durch Teufel und von Jens durch Primus.
35In der Don-Gruppe war Teufel zunächst dominierendes Weibchen. Sie bedrohte gemeinsam mit Toko außer Don alle anderen Tiere der Gruppe. Sylvia und Lena drohten zudem auch Page, Dunka und Jens, vor allem Page und Dunka.
36Der Bericht über die Fusion ist mehr oder weniger identisch mit einem gemeinsam mit Claudia Brinkmann und Cornelia Schäfer 1980 publizierten Beitrag ([254]).
37Wie bereits beschrieben war für die drei adulten Tiere, die demselben Import (B-Gruppe) entstammten, kennzeichnend die Unmöglichkeit, sie gemeinsam mit einem Weibchen, Teufel, zu halten, da sie von diesem Weibchen sofort verfolgt, gejagt und gebissen wurden. Dementsprechend drohte Teufel auch häufig aus den Nachbarkäfigen (Käfig 5, 6, 7). Die Erwin-Gruppe war ausgewogen, agonistische Interaktionen konnten innerhalb der Gruppe nicht beobachtet werden.
38Es sei hier noch bemerkt, dass nach der Separierung von Jens aus der Don-Gruppe Toko anfangs immer wieder am Zwischengitter saß und Jens Bewegungen mit den Augen verfolgte. Ebenfalls unterbrach der jetzt solitär spielende Toko, sobald Jens sich dem Käfig näherte, sogleich sein Spiel. Jens schien dieses Verhalten jedoch kaum zu beachten. Nach etwa einer Stunde gab Toko offensichtlich auf und wandte sich vornehmlich wieder dem eigenen Spiel, sowie auch dem Kampfspiel mit einem neuen Spielpartner, Don, dem stärksten Männchen der Gruppe, zu. Dies war besonders auffällig, da Don, als Jens noch in der Gruppe war, nicht spielte. Bei dem nun zu beobachtenden Kampfspiel zwischen Don und Toko zog Don Toko an Schwanz, Händen und Füßen, wobei beide den Mund weit geöffnet hatten. Weiterhin ist bemerkenswert, dass sich Teufel (B) und Toko bereits einen Tag nach Separierung von Jens (C) (im Gegensatz zu den bisherigen Beobachtungen) häufig auf dem Boden der Käfige bewegten, während Lena (C) und Sylvia (C) sich vorwiegend in den oberen Käfigbereichen aufhielten und nun auch nicht mehr bedroht wurden. Zudem fiel auf, dass Teufel nun positiven sozialen Kontakt zur benachbarten Gruppe aufnahm. So saßen Jeanette und Teufel, nur durch das Gitter getrennt, beieinander und „schmatzten“ sich zu. Auch Toko zeigte dieses Verhalten gegenüber Jeanette. An dieser Situation änderte sich bis zum 04. 01. 1978 nichts mehr.
39Agonistische Interaktionen waren nach Introduktion von Ama weitaus häufiger zu beobachten. Vor allem drohten nun Ama (B) und Sanga (B) gemeinsam Jens (C), Page (C) und Dunka (C). Diesem Drohen schloss sich, wenngleich selten, Jeanette an. Teufel (B), wieder dominierendes Weibchen, drohte hingegen selten, jedoch auch Ama (B) und Sanga (B). Den positiven und negativen Interaktionen zufolge, hatte sich die Erwin-Gruppe bereits am ersten Tag nach Amas Introduktion in zwei feste Untergruppen gespalten, nämlich in die Gruppe der stark bedrohten Tiere Dunka (C), Page (C), Jens (C), wobei Dunka und Page untereinander positiv sozialen Kontakt suchten, und in eine größere Untergruppe, bestehend aus Teufel (B), Toko, Ama (B), Sanga (B) und Jeanette. Erwin (B) nahm eine neutrale Position zwischen beiden Untergruppen ein.
40An agonistischen Aktivitäten war Lena aktiv nicht beteiligt. Sie wurde jedoch gemeinsam mit Page (C), Dunka (C) und Jens (C) von Teufel (B), Ama (B) und Sanga (B) bedroht, worauf sie sich jeweils an das Gitter zu Don und Sylvia zurückzog.
41Edith lebte bis zum 20.01.1978 in unserem Haus in Grifte ohne Kontakt zu Kapuzineraffen. Dann überführten wir sie mit ihrem Grifter Haltungskäfig in den Kapuzinerraum. Am ersten Tag saß Edith nur auf dem Käfigboden. Claudia Brinkmann berichtet über diese Introduktion in ihrer Examensarbeit. „Schon am nächsten Tag beobachteten wir, dass Edith - wie vorher - in ihrem Käfig umhersprang und sich offensichtlich nicht mehr an den anderen Tieren störte. Toko und Jeanette waren besonders interessiert an dem jungen Kapuziner. Sie versuchten, durch dass Gitter zu greifen, wohl, um mit Edith zu spielen“ (Seite 75). Edith war noch ein Flaschenkind. Von mir angebotenes festes Futter nutzte sie eher zum Spielen. In den folgenden Wochen wurde sie dann zunehmend entwöhnt. Bereits am 01.02.1978 kostete sie erstmal unseren Futterbrei, ab dem 11.02.1978 nahm sie täglich eine mit einem Esslöffel portionierte Menge an Quarkbrei zu sich, ab dem 19.02.1978 konnten wir diese Menge verdoppeln bzw. (ab dem 13.03.1978) verdreifachen. Am 16.03.1978 hängten wir für Edith in Käfig 4 einen umgebauten Schlafkasten auf, in dem wir sie ohne Risiko (wir hatten ein feinmaschiges Gitter gewählt) mit Kapuzinern vertraut machen konnten. In dem Käfig 4 hielten wir nur Sanga, die übrigen Mitglieder der Don-Gruppe waren in Käfig 3 abgetrennt. Diesen Kasten öffneten wir noch am gleichen Tag für eine Stunde. Edith kletterte vorsichtig auf diesem herum. „Nach einer halben Stunde kletterte Edith erstmalig vorsichtig am Gitter herunter auf den Boden, lief hier ausschließlich biped umher, ein Verhalten, welches ich (Claudia Brinkmann) zwar bei anderen Kapuzinern auch beobachten konnte, jedoch keineswegs mit dieser Häufigkeit. ... Edith zeigte in dieser ersten Stunde nicht das sonst zu beobachtende Fressverhalten der Kapuziner, die gebotenen Nahrungsstücke in die Hand zu nehmen, damit in höhere Käfigbereiche steigen und sie dort unter Zuhilfenahme der Hände zu fressen. Sie biss dagegen von der am Boden liegenden Nahrung ein Stück ab und lief dann kauend weiter im Käfig herum. In dieser Stunde duldete Edith auch keine Berührungen durch andere Kapuziner. Sie schrie dann laut auf und sprang entsetzt davon. Dieses Verhalten zeigte sie mehrfach gegenüber Toko, der Edith häufiger durchs Gitter spielerisch am Schwanz zog“ (Seite 76-77). Claudia Brinkmann berichtet, dass Edith in den nächsten Tagen sich sicherer und auch quadruped bewegte, sie sprang zwar schnell weg (in ihren Kasten), wenn Toko sie berührte, schrie aber nicht mehr. Auch beobachtete sie, dass Sanga und Don (aus dem Nachbarkäfig) Menschen bedrohten, versuchten diese, mit Edith zu interagieren. Am 21.03.1978 ließen wir Sangas Tochter Jeanette dazu. Edith hatte vor dieser offensichtlich keine Angst, folgte ihr sogar, soziale Kontakte konnten nicht beobachtet werden. Nämliches galt nach der Introduktion weiterer Kapuzineraffen. (Wir ließen noch am Abend des gleichen Tages Teufel mit ihrer 24 Tage alten Tochter und in den folgenden Tagen Don und Dunka hinzu.) „Nach mehr als einmonatiger gemeinsamer Haltung mit Kapuzinern wechselte Edith erstmalig ohne fremde Hilfe unter dem Käfiggitter durch zur Restgruppe. Von nun an wurde dieser Wechsel häufiger beobachtet. Allerdings blieb Edith auch im anderen Käfig ohne Sozialkontakte. Um zu klären, ob Edith besondere Beziehungen zu anderen Kapuzinern aufgebaut und ein Gruppenbewusstsein entwickelt hatte, überführten wir Don, Teufel (mit Isis), Sanga, Dunka und Jeanette wieder in den Käfig 3 zurück und sperrten Erwin, Jens, Toko, Page und Lena in Käfig 4 ab. Edith hatte, wie bereits erwähnt, die Möglichkeit, zwischen den Käfigen 3 und 4 zu wählen und entschied sich wohl unabhängig von den anderen Artgenossen für Käfig 4. ... (Für Edith) dürfte die vertrautere Umgebung attraktiver gewesen sein als vertrautere Artgenossen. Gleich am ersten Tag wurde positives Sozialverhalten Edith gegenüber deutlich, so ließ sich Edith von Erwin ohne Gegenwehr betasten. Am dritten Tag nach dem Käfigwechsel zeigten Toko und Edith zum ersten Mal Spielverhalten. Von nun an häufte sich das Spielverhalten, wobei vornehmlich Kampfspiele zu beobachten waren. Auffälligerweise spielte Edith ausschließlich mit männlichen Artgenossen und präferierte dabei das jüngste Männchen Toko eindeutig vor Jens und Erwin“ (Seite 78). Wir setzten dann Toko am 11.04.1978 in die Don-Gruppe zurück. Edith blieb in Käfig 4 und spielte solitär, besuchte aber regelmäßig Toko im Nachbarkäfig. „Bei einem dieser Wechsel zur Don-Gruppe am 13.04.1978 konnte beobachtet werden, wie Edith sich zunehmend den neuen Käfig erschloss: Der Verbindungsgang zum Außengehege war für die Don-Gruppe geöffnet, Teufel und Toko saßen in Käfig 3 nebeneinander. Dazu kam Edith und sogleich spielten Edith und Toko miteinander. Als nach einer Weile Teufel aufstand und in den Außenkäfig lief, folgte ihr Toko. Edith lief Toko nach, blieb aber an der Öffnung des Verbindungsganges zurück, um dann, wenn Toko mit Teufel in den Innenkäfig zurückkam, das Spiel wieder aufzunehmen. Dieses wiederholte sich dreimal. Zunehmend folgte Edith auch in den Gang, ging aber nie ins Freie und kehrte sofort zurück, sobald ein anderes Tier von den Außenkäfigen aus den Gang betrat. Als Toko das vierte Mal das Spiel unterbrochen hatte, ..., folgte nun auch Edith zögernd durch den Gang ins Freie, wo sie sich nur kurz und äußerst vorsichtig bewegte und auch draußen ohne Kontakt zu Toko blieb“ (Seite 79).
42In der Tabelle habe ich zwei Namen in Klammern angegeben. Jonas war kein Junge, er erhielt Jahre später den Namen Jonaine (s. u.). Teufel war nur die biologische Großmutter von Ediths Kindern.
43Bubi wurde nicht schwer verletzt, war aber ständiges Ziel von Erichs Attacken und schien „psychisch“ nicht den Verlust der dominierenden Position zu ertragen. Seine physische Konstitution wandelte sich dramatisch, ein fatales Ende konnte erwartet werden. Daher separierten wir ihn und hielten ihn allein ([248]).
44Das abendliche Einsperren der Kapuzineraffen gestaltete sich für mich persönlich immer schwieriger. Wir mussten zum Einsperren den Außenkäfig betreten und den Durchgang nach Innen mit einer Klappe schließen. Dies war für mich eigentlich problemlos zu erledigen (meine Mitarbeiter benötigten teilweise einen Wasserschlauch, um den Affen die Dringlichkeit des Einsperrens nahezubringen.) Dies änderte sich aber schlagartig nach Bubis Reintroduktion in die Hoppediz-Gruppe. Sobald Bubi mitbekam, dass ich diese Gruppe einsperren wollte, raste er heraus, setzte sich auf einen meiner Schuhe und drohte massiv Hoppediz. So verbrachte ich überredend manchmal viel zu viel Zeit vor der Schieberklappe, von innen drohte Hoppediz massiv und schlug gegen die Klappe, von außen Bubi, und ich stand da, jederzeit damit rechnend, dass Hoppediz mich beißen würde. Hierdurch bedingt war dann auch Hoppediz der einzige Kapuzineraffe, dem ich bei dem Abendrundgang nichts zustecken konnte. Einmal steckte ich ihm ein Futterpellet in den Mund, sagte: „So schlimm bist du gar nicht!“und machte den Fehler, ihm noch ein weiteres Stück geben zu wollen. Hoppediz erinnerte sich an seine Feindschaft und biss zu. (Mein Finger hat dies überstanden).
45Bei der handaufgezogenen Edith protokollierte ich bereits am vierten Lebenstag das Sichkratzen mit einer Hinterextremität, am elften Lebenstag das Sichkratzen mit der linken Hand. Klammerte sie sich anfänglich fast bewegungslos an ihren Fellhandschuh, kletterte sie bereits am achten Lebenstag aus dem Korb und am zwölften Lebenstag auf meine Hand. (Mit ihrem Rollschwanz konnte sie sich bereits am fünften Lebenstag am menschlichen Finger festhalten.)
46Edith kletterte am 48. Lebenstag erstmals aus ihren Käfig und am 62. Lebenstag auch vom Gitter auf meinen Schoß.
47Edith spielte in ihrem dritten Lebensmonat mit uns und mit Gegenständen und lief am 75. Lebenstag erstmals auf dem Boden des Zimmers herum.
48Im vierten Lebensmonat kletterte Edith auf unsere Schultern und aß erstmals festes Futter (Banane) aus dem Mund meiner Frau. (An angebotener Nahrung hatte sie kein Interesse.) In diesem Monat begann sie zu springen und überbrückte Distanzen von 60 cm dabei mühelos.)
49Insofern sind die engen sozialen Beziehungen der drei dominierenden Männchen in der Don-Gruppe, über die ich noch berichten werde, besonders hervorzuheben.
50Eines Morgens saß ich in der Cafeteria mit meinen Mitarbeitern, da setzte sich ein mir bis dahin nicht bekannter Mathematiker an unseren Tisch und sagte: „Wir nennen Sie nur noch den Affenzauberer“. Wie kam ich zu dieser Ehre? Die Außenanlagen auf dem Werkstattgebäude waren von den gegenüberlegenden Büroräumen gut einsehbar. So konnten - von mir unbemerkt - zahlreiche Zuschauer folgende Episode beobachten. Bubi war meinen Tierpflegern entwischt. Verzweifelt und erfolglos versuchten diese ihn in seinen Käfig zurückzudrängen. (Bubi reagierte bei Bedrängungen jeglicher Art mit Angriff.) Schließlich riefen sie mich an und baten um Hilfe. Natürlich eilte ich herbei, betrat das Außengelände, hob meine Arme und rief „Bubi“, und Bubi rannte - so schnell er konnte - laut schreiend auf mich zu und folgte mir in das Außengehege zurück. Die mittlerweile zahlreichen Beobachter meinten wohl, ich könnte so Affen einfangen, dies konnte ich aber nur mit Bubi. Keine Zauberei, vielmehr echte Zuneigung.
51Erst in diesen Tagen ist mir eine Arbeit von Hediger und Zweifel ([69]) beim Sortieren von Separata in die Hände gefallen, in der nämliches von Pfyfer, einem 36 Jahre alten Kapuzinermann, berichtet wird.
52Den beiden Studentinnen gefielen auch andere Affen, sie schrieben ihre Staatsexamensarbeiten über Springaffen (vgl. Kapitel 8).
53Kim trug ein Neugeborenes, das wir nicht riskieren wollten.
54Die Männchen Tschiggo und Primus und die Weibchen Purzel und Inge wurden aus der Don-Gruppe verdrängt, zwei Weibchen (Ama, Baja) waren gestorben. Zusätzlich zu den vier verdrängten Individuen hatten wir in Abänderung meiner Haltungsstrategie, möglichst keine unnötigen Veränderungen vorzunehmen, Lena aus der Don-Gruppe entnommen. Unterstützt von ihrer „Freundin“ Sylvia bedrohte sie so massiv alle anderen Gruppenmitglieder, so dass wir uns entscheiden mussten, entweder die Don-Gruppe wieder aufzulösen oder Lena zu entfernen. Ich habe mich gegen Lena entschieden. Alle fünf Kapuzineraffen konnten in die Bubi-Gruppe (re-)introduziert werden. Lena, die vorher in dieser Gruppe nicht gelebt hatte, musste sich mit einer peripheren Position abfinden.
55Petra Pippert diskutierte in ihrer Dissertation das Theoriengebäude der Soziobiologie und prüfte auch Literaturbefunde zum „paternal behaviour“ bei anderen Primatenspecies, worauf ich hier nicht eingehe.
56Auch bei entsprechenden Untersuchungen an unserer eigenen Species werden Vaterschaften ausgeschlossen und nicht belegt.
57Nur Jungtiere, deren Mütter keine eindeutigen Präferenzen für ein bestimmtes Männchen aufzeigen, orientieren sich anders.
58Diese Aussage gilt meiner Meinung nach zumindest für alle nichtmenschlichen Primaten. Sie stützen nicht das Theoriegebäude der Soziobiologie. Gesteuert von ihren Genen müssten Individuen folgendes leisten: Sie müssten grundsätzlich eigensüchtig handeln und nur auf ihren eigenen Fortpflanzungserfolg bedacht sein (Darwinian fitness); Handeln sie altruistisch, dann müssten sie zwangsläufig den Verwandtschaftsgrad des jeweiligen Partners bei ihren Investitionen einrechnen (Inclusive fitness); zudem muss konsequenterweise ein Individuum - gesteuert von seinen Genen - genau wissen, ob der Partner mit ihm verwandt ist oder nicht. Kapuzineraffen - und auch andere Primaten - verhalten sich „falsch“, sie präferieren Vertrautere vor weniger Vertrauten, ohne auf die tatsächliche Verwandtschaft zu achten ([247]).
59In der Don-Gruppe dominierten zwei mächtige Clane, der Sylvia- und der Teufelclan. Nach Don war Teufels Sohn Toko das mächtigste Männchen. Dunka hatte eine Randposition. 1988 verbündeten sich ihre drei adulten bzw. subadulten Söhne, Daniel (1978), Dennis (1980) und David (1982), gegen Toko und griffen diesen gemeinsam an. Gegen drei große Männchen war selbst Toko chancenlos. Gleichzeitig griff Dunka die anderen Weibchen an. Ich entschloss mich, Dunka und ihre Söhne aus der Gruppe zu entfernen. Wir hielten diese separat. Aus unseren Haltungserfahrungen wussten wir, dass Mütter als Sexualpartner für ihre Söhne unattraktiv sind. Dieses Wissen wurde relativiert, Dunka gebar am 14.04.89 Doria, Vater konnte nur einer ihrer Söhne sein.
60Große Sozialverbände gesellig lebender Primaten werden kontinuierlich in der Regel nur an der von Menschen angelegten Futterstelle beobachtet. Die Beobachter füttern ihre Tiere an oder beobachten sie in der Nähe von Tempelanlagen, wo die Affen aus religiösen Gründen von der einheimischen Bevölkerung gefüttert werden. Dabei achten verständlicherweise sowohl die Beobachter als auch die lokale Bevölkerung darauf, dass die Affen nicht darben.
61Für Affen ist generell ein Gitter eine Bereicherung des Lebensraumes.
62Auch Hoppediz konnte nicht lesen. Insofern folgten auch hier die Kapuzineraffen nicht den Annahmen der Soziobiologie. Für das Töten dürfte die soziale Situation in der Gruppe verantwortlich gewesen sein. Zweifellos hatte das dominierende Männchen in dieser Gruppe keine unbestrittene Position, wurde er doch von den Weibchen nicht als der attraktive Mann angesehen. Fragen wir uns, warum ein nicht eindeutig dominierendes Männchen Kinder töten könnte, liegt die Erklärung wohl an dem Verhalten der Weibchen. In Kapuzineraffengruppen sind Kinder tragende Weibchen attraktiv und ändern ihr Verhalten. Selbst Weibchen, die vor der Geburt peripher lebten und bedroht wurden, sind nun im Zentrum der Sozialgruppe und werden - anthropomorph betrachtet - selbstsicherer. Diese verbesserte soziale Position verlieren sie aber nach der Entwöhnung des Jungtieres. Sie müssen sich mit ihrer alten sozialen Stellung zufrieden geben. Insofern ändert das Töten des Kindes die Stellung der Mutter, gleichzeitig verbessert sich relativ die Position des Töters. Dabei scheint es unwichtig zu sein, ob das Kind sein eigenes oder das eines anderen Männchens ist. ([248])
63Einmal beobachtete ich einen Konflikt zwischen Don und Toko, beide drohten in die gleiche Richtung und schlugen die Köpfe gegeneinander ohne direkten Blickkontakt.
64Die Beobachter protokollierten das Verhalten nach einem vorgegebenen Katalog.
65In der Sylvia-Untergruppe sind auch die aktiven Putzkontakte der Sarah-Tochter Silke zu ihren Onkeln Samuel (n = 29) und Philipp (n = 16) und ihren Cousinen Tonia (n = 19) und Paula (n = 21) erwähnenswert. Sarahs Tochter Franziska putzte ihre Schwester Stephanie (n = 18) und ihre Nichten Tonia (n = 10) und Paula (n = 6). Petras Tochter Pia schließlich putzte ihre ein Jahr ältere Tante Sandra (n = 11) und ihre Schwester Paula (n = 16).
66Teufel putzte aktiv nur ihre Tochter Jesca (n = 12), wurde aber außer von Jesca (n = 62) auch von ihrer Tochter Isabelle (n = 30) und ihrem Sohn Jim(n = 19) geputzt. Isabelle unterhielt gegenseitige Putzbeziehungen zu ihrer Schwester Jesca (aktiv n = 28, passiv n = 16) und putzte ihren Neffen Jan (n = 10) und ihren Sohn Miratus (n = 32), sie selber wurde von ihrem Bruder Justus (n = 6) geputzt. Jesca putzte zudem ihren Bruder Jim (n = 6) und ihre Neffen Jan (n = 20) und Miratus (n = 7). Jim schließlich putzte seinen Bruder Justus (n = 38), der zur Untergruppe um Kim und ihrer Tochter Karin vermittelte.
67Edith putzte ihre Söhne Edgar (n = 19) und Elvis (n = 34).
68Claudia putzte ihre Söhne Cris (n = 6) und Clarence (n = 14) und ihre Tochter Carmen (n = 35) und wurde von ihrer Tochter Casparia (n = 23) geputzt.
69Ihre Mutter Dunka hatten wir gemeinsam mit ihren Brüdern separiert (s. o).
7016.05.1992 Kurtisane (Kim), 01.06.1992 Isidor (Isabelle), 02.06.1992 Sunny (Sarah), 19.06.1992 Jolan (Jesca) und am 01.07.1992 Samson (Stephanie).
71Von dieser Chance, das Zusammenleben einer etwa vierzigköpfigen Kapuzineraffengruppe beobachten zu können, waren Gäste der Primatenstation begeistert. Nur einmal erlebte ich eine andere Situation. Ich hatte sogenannte Tierschützer eingeladen, ich wollte ihnen die Möglichkeit einräumen, in einem von mir angebotenen Kolloquium zum Thema „Artenschutz, Tierschutz, Tierversuche“ ihre Positionen zu vertreten. Bei unserem Vorgespräch im Beobachtungsraum hielt sich eine Dame des Tierschutzvereins ständig die Augen zu. Besorgt fragte ich diese, ob sie Augenprobleme hätte, und bekam die Antwort, sie könne das Leid (aktive Kapuzineraffen) nicht ertragen.
72Bei diesen Versuchen erfüllte ein ungeheurer Lärm den an sich schon lauten Kapuzinerraum, fast alle Kapuzineraffen, besonders die männlichen, belegten mit ihrem angeborenen Verhalten, wieso die Kapuzineraffen in Brasilien „die Hämmerer des Waldes“ genannt werden.
73Wir werteten für diese Befunde die Daten von vierzehn bei uns geborenen Kindern aus, die Daten von acht dieser Kinder beruhen auf Einzeltierbeobachtungen an fünf männlichen und drei weiblichen Kindern. Auf diesen Beobachtungen beruhen auch die angegebenen quantitativen Befunde ([253]).
74Dabei bauen sie mehr oder wenger zwangsläufig auch Beziehungen zu deren Müttern auf und dürfen auch an diesen saugen.
75Dabei handelt es sich - wie bereits betont - jeweils in der Regel nur um ein bestimmtes Männchen.
76Im Hinblick auf spätere auffällige Unterschiede zwischen den Geschlechtern sollen erste - nicht gesicherte - Unterschiede nicht verschwiegen werden:
Adulte Männchen und Weibchen bevorzugen bei dem Gehenzu, dem Zusammensitzen und dem Spiel männliche Kinder, weibliche Kinder dagegen werden häufiger geputzt.
Geschwister sind für männliche Kinder attraktiver als für weibliche. Dementsprechend sitzen Geschwister häufiger mit männlichen Kindern zusammen und spielen mit ihnen als mit weiblichen Kindern. Weibliche Kinder werden aber häufiger von Geschwistern aufgesucht, manipuliert und sind häufiger Partner bei dem Schmusen. Nämliches gilt für im gleichen Jahr geborene Jungtiere, doch sind hier die Unterschiede noch geringer.
Bei den generell nur selten zu beobachtenden Kontaktaufnahmen anderer nicht verwandter Juveniler ist zudem auffällig, dass männliche Juvenile männliche Kinder und weibliche Juvenile weibliche Kinder präferieren.
77Wir werteten für diese Befunde die Daten von 15 bei uns geborenen Kindern aus, acht Männchen und sieben Weibchen ([252]).
78Zudem soll erwähnt werden, dass Töchter ihre Mutter signifikant häufiger manipulieren als Söhne.
79Wir protokollierten das aktive Gehenzu 1290 ± 141 mal für männliche und 1147 ± 381 mal für weibliche Kinder, das passive Gehenzu 1460 ± 264 bzw. 1235 ± 357 mal.
80Wir protokollierten das aktive Manipulieren 47 ± 6 mal für männliche und 75 ± 47 mal für weibliche Kinder, das passive Manipulieren 56 ± 12 bzw. 64 ± 40 mal.
81Wir protokollierten das Kontaktsitzen 911 ± 85 mal für männliche und 994 ± 117 mal für weibliche Kinder.
82Wir protokollierten das Schmusen 81 ± 16 mal für männliche und 56 ± 29 mal für weibliche Kinder.
83Wir protokollierten das aktive Putzen 9 ± 2 mal für männliche und 8 ± 5 mal für weibliche Kinder, das passive Putzen hingegen 195 ± 53 bzw. 80 ± 38 mal.
84Für diese Untersuchung analysierten wir die Befunde von 21 Kapuzineraffenkindern, die in der Don-Gruppe heranwuchsen. Drei dieser Kinder wurden über die gesamten vier Lebensjahre beobachtet, die anderen ein bis drei Jahre lang. Die Ergebnisse des zweiten bis vierten Lebensjahres beruhen auf Beobachtungen an jeweils 14, die des fünften Lebensjahres beruhen auf solchen an 10 Individuen ([257]).
85Für diese Untersuchung standen die Daten von 18 juvenilen Kapuzineraffen, neun Männchen und neun Weibchen, die alle in der Don-Gruppe aufwuchsen, zur Verfügung. Die Don-Gruppe bestand zum Zeitpunkt der Befundaufnahmen aus bis zu 43 Individuen. Die 18 untersuchten Individuen erlaubten die Analyse von 53 dyadischen Beziehungen - zwölf Bruder-Bruder-Dyaden, 15 Schwester-Schwester-Dyaden und 26 Bruder-Schwester-Dyaden. Wir errechneten wieder einen Erwartungswert, bezogen diesen aber auf alle möglichen Sozialpartner mit Ausnahme der Mutter ([258]).
86Hella Kröger
87Hella Kröger verglich die mittlere Häufigkeit mit dem Erwartungswert, dem Wert, der zu erwarten wäre, wenn alle Gruppenmitglieder gleichhäufig mit dem jeweiligen Kapuzinerkind interagieren würden. Bei der Kalkulation des Erwartungswertes für die (neben der Mutter) übrigen Gruppenmitglieder wurden die Kontakte zur Mutter nicht berücksichtigt.
88Ich hätte diese Bebachtungen auch als Nachweis des Erkennens der väterlichen Verwandtschaft publizieren können, habe es aber vorgezogen, diese Befunde auf Kontakte zu Individuen des gleichen Altersbereiches zurückzuführen, und einer Publikation widerstanden.
89Bereits mehrfach habe ich in diesem Buch auf die Schwierigkeiten der Geschlechtsbestimmung hingewiesen. Meine Mitarbeiterin Petra Pippert hatte dem kleinen Kapuzineraffenkind den Namen Jonas gegeben. Bei mir und keinem der späteren Beobachter tauchten Zweifel auf. Auch Elisabetta Visalberghi, die monatelang unsere Kapuzineraffen beobachtet hatte, fiel dieser Fehler nicht auf. Nach der Feststellung des richtigen Geschlechtes schrieb ich ihr nach Rom und bekam die Antwort, sie müsse zwar nun die Datenauswertung überarbeiten, doch würden die Ergebnisse besser. Auch bei ihren Experimenten hatte sich Jonas weniger am Objektspiel beteiligt als andere „Männchen“. Ich hatte mich damals über diesen Irrtum gefreut, konnte ich doch so die Validität meines Beobachterteams unbeabsichtigt im Nachhinein testen. Eigentlich sind die Unterschiede zwischen den Geschlechtern gut erkennbar, beide haben ein erigierbares Geschlechtsorgan, doch ist dies bei Weibchen eine Rinne (Abbildung 13.39), bei den Männchen eher ein Stempel, was man bei Tschiggo (Abbildung 13.41) erkennen mag.
90Die quantitativen Daten erlauben jedoch nicht, nach der Häufigkeit des Kontaktsitzens auf geschlechtstypisches Verhalten zu schließen. Jüngere Männchen sitzen teilweise häufiger in engem Körperkontakt mit anderen Individuen als jüngere Weibchen. Hierfür ist die Zusammensetzung der jeweiligen Familiengruppe verantwortlich. ([255])
91Kapuzineraffen rütteln angeborenermaßen an allem Rüttelbaren und erschließen sich so im Freiland neue Ressourcen bzw. verjüngen - wie ein Bäume beschneidender Gärtner - die Bäume ihres Lebensraumes.
92Dies war nur bei den in größeren Gruppen lebenden Affen notwendig, da bei diesen das gesamte Gehege und vor allem die stark verschmutzten Ruhebretter täglich sorgfältig gereinigt werden mussten.
93Michael Dulitz beobachtete diese Gruppe bei der Befunderhebung für seine Staatsexamensarbeit vor der Geburt von Purzels Sohn Plato über 99 Tage. Während seiner Beobachtungen Oktober bis Dezember 1994 lebten noch das Wildfangweibchen Migga und seine 1979 geborene Tochter Dido im Sozialverband, Elka, Eva und Plato waren noch nicht geboren. Er berichtet zum agonistischen Verhalten: „Insgesamt fällt auf, dass agonistische Verhaltensweisen innerhalb der Kapuzinergruppe weit seltener als positive Interaktionen auftreten. ... Das Drohen der Männchen ist überwiegend gegen Mitglieder der benachbarten Don-Gruppe gerichtet. ... Vor allem das Weibchen Purzel ... wird überdurchschnittlich häufig bedroht. Fälle von agonistischem Zerren, Jagen oder Beißen wurden zwischen Mitgliedern der Hoppediz-Gruppe insgesamt 40mal von mir protokolliert. 22 dieser Aktionen richteten sich allein gegen Purzel, darunter alle sechs Fälle von Beißen, die ich überhaupt innerhalb der Gruppe protokolliert habe (Seiten 8-9)“.
94Ihre Schwester Alice war am 25.11.1996 gestorben.
95„So ist besonders auffällig, dass Plato von seiner Mutter sehr viel häufiger getragen wurde als das gleichalte weibliche Kind Eusebias. ... Aufgrund ihrer Außenseiterposition war Plato für (Purzel) die Chance, mehr Sozialkontakte, insbesondere mit den Weibchen des Erna-Clans zu erhalten. ... Daher war es für sie von Vorteil, Plato so häufig und solange es irgend geht, zu tragen, denn die Vermutung liegt nahe, dass mit der wachsenden Selbstandigkeit Platos die Kontakte zu den anderen Tieren im gleichen Maße wieder abnehmen werden. Vielleicht stellt Plato fur sie auch eine Art „Schutz“ vor agonistischen Attacken gegen sie dar“ (Seiten 68-69).