Abbildung 14.1: Nikita
Ich ließ beide Raumtüren offen, öffnete die Tür des Haltungskäfigs (und alle Türen in
den Gehegen des neuen Makakenraumes), meine Mitarbeiterin Barbara Lührmann und
ich warteten - „verborgen“ in der Futterküche -, doch wurde unser Warten nicht
belohnt, unsere fünf Javanermakaken (Nikita, Vanda nebst Frieda und Vera nebst
Paula) blieben in ihrem geöffneten Gehege. Sie hatten sozusagen „den Braten
gerochen“, sie wollten ihr sicheres Gehege nicht verlassen. Wir mussten sie erst
mühselig aus ihrem Haltungsgehege jagen, dessen Tür verschließen und dann sie
dringend auffordern, den Kapuzinerraum zu verlassen und den nun fertigen
Makakenraum aufzusuchen, was sich stundenlang hinzog. Einmal auf den Flur
getrieben, suchten sie dann den neuen Raum und die neuen Käfige sofort
auf.
Am 21.05.1975 importierte Werner Meinel weitere fünfzehn Weibchen, eines war stark
verletzt und wurde für anatomische Untersuchungen entnommen, zwei Weibchen
starben während der Quarantäne, so dass nun insgesamt 17 Javanermakaken in dem
Makakenraum lebten. Einige der Weibchen waren bei Import gravid, doch im
offensichtlichen Gegensatz zu den Kapuzineraffen (Kapitel 13), bei denen
schwangere Weibchen ihre Jungen auch austrugen, abortierten alle Weibchen
(und aßen die Föten auch auf). Dies ist besonders bemerkenswert, da in den
folgenden Jahren der Haltung die Anzahl von Aborten vernachlässigbar war. Es
muss dahingestellt bleiben, ob der Transportstress oder aber die Anwesenheit
eines Männchens (Nikita) im Haltungsraum für das Abortieren verantwortlich
war.
Der „Versuchsaffe“ in der Forschung war damals der Rhesusaffe Macaca
mulatta, sogenannte „Affenergebnisse“ also Ergebnisse an Rhesusaffen
(Abbildungen 14.9 - 14.12). Wegen der zunehmendem Exportrestriktionen in
Indien2,
dem Hauptexportland wurde dieser in Europa zunehmend durch Javanermakaken
ersetzt. Ergebnisse an Javanermakaken stimmten mit denen an Rhesusaffen überein,
dies gilt auch für ihr Verhalten. Japanische Kollegen erheben ihre Befunde vornehmlich
an Japanmakaken Macaca fuscata (Abbildungen 14.13 - 14.14). Zudem stehen bzw.
standen in amerikanischen Primatenzentren auch Kolonien des Schweinsaffen
Macaca nemestrina (Abb. 14.91) und des Indische Hutaffen Macaca radiata
(Abbildungen 14.15 - 14.16) als Forschungsobjekte zur Verfügung. Auch bei
Untersuchungen von Makaken kommt der genauen Artbestimmung Bedeutung
zu.3
Der einzige nichtasiatische Makake, der nordafrikanische - und auch auf Gibraltar vorkommende - Berberaffe Macaca sylvana (Abbildungen 14.17 - 14.19) hat als „Versuchaffe“ kaum Verwendung gefunden. Er diente nur als Beobachtungsobjekt für Verhaltensbeobachtungen. Nach seinem Verhalten ist er auch nur begrenzt mit den übrigen Arten des Genus Macaca vergleichbar, nutzen doch bei dieser Art die Männchen die Jungtiere sozusagen als Schutzschild, um ungefährdet mit anderen Männchen zu interagieren (Abb. 14.19). Diese Arten stelle ich mit den folgenden Abbildungen vor. Die meisten Untersucher des Makakenverhaltens bevorzugten Orte, an denen ihre Versuchsobjekte gut beobachtbar sind. Wie ich bereits an verschiedenen Stellen in diesem Buch berichtet habe, ist das Beobachten und das Finden der Affen im Blätterwald nicht einfach. Auch unsere intensive und letzlich erfolglose Suche nach Mohrenmakaken (Macaca maura) habe ich nicht verschwiegen (Kapitel 8) habe ich nicht verschwiegen. Anschließend suchten meine Frau und ich im Süden Sulawesis weiter und waren erfolgreich (Abbildungen 14.4 - 14.5). Verhalten konnten wir freilich nur sehr begrenzt beobachten. Freilebende Javanermakaken trafen wir auf Bali. Auf Bali konnten wir zudem Javanermakaken unter sogenannten halbnatürlichen Bedingungen beobachten. Ich riet meiner Frau, ihre Brille abzunehmen, doch hörte sie nicht auf mich. Die „frei“ lebenden Makaken waren trainierte Diebe.


Wir hatten also mit unseren Javanermakaken „typische“ Versuchsaffen zur Verfügung.
Ein ethologisches Forschungsinteresse meinerseits bestand nicht, schienen
doch Makaken „ausgeforscht“, zudem war ich an dem langfristigen sozialen
Zusammenleben in stabilen Sozialgruppen interessiert und nicht an kurzfristigen
Beobachtungen.4
Dann hörte ich einen Vortrag des Züricher Primatologen Hans Kummer, der
Zweierbeziehungen bei Blutbrustpavianen (Therpithecus gelada) untersucht und ein
Stufenmodell der zunehmenden Integration vorgestellt hatte ([117]). Dieses fand ich
hochinteressant und bat Werner Meinel um die Erlaubnis, mit seinen Versuchstieren
vor deren „Verbrauch“ entsprechende kurzfristige Untersuchungen durchführen zu
dürfen.
Dass diese Beobachtungen zu so interessanten Forschungsergebnissen führen sollten, hatte
ich damals nicht erwartet. Über mehr als zwei Jahrzehnte habe ich dann an Javanermakaken
gearbeitet.5
In den diesem Kapitel zugrunde liegenden Publikationen belegen zahlreiche
Abbildungen die Befunde, auch habe ich in diesen die umfängliche Literatur über
Makaken sorgfältig dokumentiert und diskutiert. Den unzähligen Autoren sei hier
ausdrücklich gedankt. Sie haben präzise Fragen aufgeworfen, die wir experimentell mit
Hilfe unserer Makaken-Individuen beantworten konnten.
Durch die kontinuierliche Förderung durch die Deutsche Forschungsgemeinschaft
konnte ich viele offene Fragen experimentell klären, unterstützt
durch zahlreiche studentische und auch wissenschaftliche Mitarbeiter.
Barbara Lührmann ([126]), Cornelia Schäfer-Witt ([194]) und Annette
Klaiber-Schuh6
schrieben über diese Art ihre Dissertationen. An der Arbeit wirkten zudem mit:
Heribert Arend, Gesine Boehlke, Rita Engelhardt, Meike Erbarth-Fischer,
Christiane Fiege, Martina Grebe, Elke Harigel, Regina Hennek, Beate
Hollstein, Ursula Lingelbach, Gerhard Neuhoff, Ingrid Pampuch, Manfred
Paul, Lieselotte Schindler, Petra Schroer, Elke Sobisch und Christiane Möller.
Zahlreiche studentische Mitarbeiter verfassten auch über Teilergebnisse
der Langzeitstudie ihre Examensarbeitenen, die mir leider nur teilweise
vorliegen.7
Unser Hauptforschungsinteresse galt dem Phänomen der Rangordnung und den
unterschiedlichen Persönlichkeiten in der Makakengesellschaft. Parallel hierzu
untersuchten wir die auffälligen Geschlechtsunterschiede im Verhalten, die ich in
diesem Buch bereits für Totenkopfaffen (Kapitel 11) und Kapuzineraffen (Kapitel 13)
dokumentiert habe. Diese galten nach dem Stand der Literatur für alle Arten des
Genus Macaca. Sicherlich auch Zeitgeist bedingt, wurde in den Arbeiten die Frage
nach den Ursachen dieser Unterschiede gestellt. Da die jungen Makaken je nach
Geschlecht unterschiedlich von ihren Sozialpartnern behandelt werden, schien die
Hypothese erlaubt, dass hierfür möglicherweise Erziehungsprozesse verantwortlich sind.
Dies konnten wir eindeutig falsifizieren, wir erhoben umfangreiches Datenmaterial an
in der Gruppe geborenen (Unterkapitel 14.5) und an handaufgezogenen (Unterkapitel
14.6) Individuen. Auch hierüber werde ich noch ausführlich berichten (Unterkapitel
14.5, 14.6).
Es ist mir noch ein Bedürfnis dem vielleicht nach dem Lesen dieses Kapitels
entstehenden Eindruck, Makaken seien besonders aggressiv, entgegen zu treten.
Das rigide Dominanzsystem der Makaken (und wohl aller meerkatzenartigen
Altweltaffen) dient vor allem der Vermeidung von Konflikten im Sozialverband. In der
Evolutionsgeschichte dürften entsprechende Sozialsysteme sich unter jahreszeitlich
kargen Bedingungen entwickelt haben. Sie gewährleisten, dass zumindest der
höherrangige Teil der Population, der alle Vorrechte hat, überlebt, was zwangsläufig
für die niederrangigen Individuen fatale Folgen hat. Wann in der Millionen Jahre
langen Evolutionsgeschichte sich ein solches Verhalten als selektionsbegünstigt
herausgestellt hat, können wir freilich heute nicht entscheiden. Insofern sind nach
meiner Einschätzung häufig angestellte Versuche, das ererbte Sozialsystem
mit heute vorzufindenden ökologischen Bedingungen zu begründen, nicht
weiterführend.
Bei8
Javanermakaken - wie generell bei allen Species des Genus Macaca und wohl nahezu
allen Vertretern der Cercopithecinae - beruht das Sozialsystem der Gruppe auf einer
starren hierarchischen Rangordnung, die hinreichend ist, um in der Gruppe zu
beobachtende Interaktionen zu erklären (u. a. [247]).
Es soll hier betont werden, dass gerade nach den in Kassel durchgeführten
vergleichenden Untersuchungen verschiedener Primatenspecies, die ich in den
bisherigen Kapiteln dieses Buches dokumentiert habe, das Vorhandensein einer
Rangordnung der Spezialfall innerhalb der Primates ist. In der Regel finden wir
bei den Primaten - wie bei allen anderen gesellig lebenden Species - zwar
Dominanzbeziehungen, also das banale Ergebnis, dass nicht alle Tiere der Gruppe
gleich sind, doch keine echten Rangordnungen. Dabei wollen wir von Rangordnungen
nur sprechen, wenn das Grundprinzip der sozialen Organisation ist, dass das
Ergebnis von Auseinandersetzungen beliebiger Zweierbeziehungen eindeutig
vorhergesagt werden kann, und wenn der Überlegene zudem gleichzeitig über
alle oder fast alle Individuen dominiert, die dem Unterlegenen unterlegen
sind.
Bei Species mit Rangordnungsstrukturen ist für das Individuum seine Position in der
Gruppe bestimmend. Je nachdem, ob das Individuum im oberen oder im unteren
Bereich der Hierarchie eingeordnet ist, bzw. ob es z. B. im oberen Bereich die Alpha-,
Beta- oder Gamma-Stellung hält, unterscheidet es sich auffällig von anderen Tieren
der Gruppe.
Darüber hinaus beobachten wir in Makakengruppen, dass in der Gruppe geborene
Jungtiere den Rangplatz ihrer Mütter übernehmen, sozusagen erben. Dies gilt
uneingeschränkt für weibliche Individuen, die in der Gruppe verbleiben. Unsere Daten
sprechen zudem dafür, dass Nämliches auch für männliche Individuen zutrifft, dass
also auch hier der endliche Rang durch den Rang der Mutter vorbestimmt
wird.
Fassen wir zusammen, dann ist es für das Individuum ohne Zweifel entscheidend,
welchen Rang seine Mutter besitzt, da dieser für ihn und für seine Nachkommen
geradezu schicksalhafte Bedeutung hat.
Wodurch wird nun - und dies war auch die eigentliche Frage des gesamten
Forschungsvorhabens - die Weitergabe des Rangplatzes an die Nachkommen
gewährleistet?
Beobachten wir die Interaktionen in der Gruppe, dann stellen wir fest, dass Kinder
bereits sehr früh - in der ersten Lebensphase - von ihren Müttern und Verwandten
unterstützt werden. Diese Unterstützung ist dann auch für den Rest des Lebens
permanent aufzeigbar. Durch zahlreiche Veröffentlichungen zu diesem Thema wurde
die Schlussfolgerung nahegelegt, dass diese Unterstützung für den späteren Rang des
Individuums ursächlich sei. Ein Beleg für diesen Zusammenhang stand jedoch aus, die
Kausalität ist demnach nur scheinbar.
Als wir 1974 unsere Untersuchungen zur Sozialstruktur des Javanermakaken
begannen, drängte sich uns bei der Beobachtung von Gruppenbildungsprozessen - zehn
Weibchen wurden sukzessive in eine Ein-Männchen-Gruppe überführt - der
Eindruck auf, dass es unterschiedliche Typen von Weibchen gäbe. Je nach
ihrem Verhalten in der ersten Zeit nach Introduktion nahmen sie später einen
höheren oder einen niedrigeren Rangplatz in der Gruppe ein (Unterkapitel
14.2).
Die Integration jedes einzelnen Weibchens verlief grundsätzlich nach dem gleichen
Muster, so wurde es von dem einzigen Männchen solange verfolgt und gebissen, bis es
keinen Widerstand mehr leistete. Die genauere Analyse erbrachte, dass zwischen
Männchen und neuem Weibchen bestimmte Prozesse ablaufen müssen, bevor von
einem Abschluss der Integration gesprochen werden kann: Die Integration verläuft
über verschiedene Stadien, die jeweils obligatorisch sind. Insgesamt konnten fünf
verschiedene Stadien unterschieden werden, so folgte auf das (1) Verfolgen durch das
Männchen das (2) Präsentieren durch das Weibchen, das (3) Besteigen und ein
rituelles (4) Beißen in den Rücken, erst danach konnte als letztes Stadium die (5)
gegenseitige soziale Körperpflege beobachtet werden. Unterschiedlich war nun die Zeit,
die die jeweiligen Weibchen benötigten, um das Stadium 5 zu erreichen. Einige
schafften dies bereits am Tag der Introduktion, andere benötigten hierzu mehrere
Monate.
Im Anschluss an ihre Integration mussten sich neu introduzierte Weibchen mit dem
untersten Platz in der Hierarchie zufrieden geben. Sie suchten durchgängig
den engen Sozialkontakt zum jeweils Rangnächsten, verhinderten dadurch
gegen sich gerichtete agonistische Aktivität und wurden mehr oder weniger
gleichrangig. Dann stellten sie besonders enge soziale Beziehungen zu dem
nächsthöheren Individuum her, griffen dann gemeinsam mit diesem den ehemaligen
Rangnachbar an und wurden so höherrangiger als der ehemalige Rangnachbar. Über
diese Strategie verbesserten einige Individuen ihren Rang Schritt für Schritt,
andere hingegen mussten mit einem niedrigeren Rang vorliebnehmen, bzw.
verloren - trotz bester Vorbedingungen (frühere Introduktion) - ihren Rangplatz
sukzessive.
Diese Beobachtungen und weitere Kontrollexperimente waren die Basis für das
weitere Vorgehen. Wir wollten prüfen, ob der gewonnene Eindruck, es gäbe
a priori unterschiedliche Typen von Weibchen, belegt werden könnte. Eine
Alternativerklärung wäre nämlich gewesen, dass die beobacheten Ränge nur zufällig
waren, abhängig von Beziehungen der Individuen untereinander. So könnte die
Freundschaft9
zum Apha-Weibchen z. B. einen hohen Rang bedingen, seine Freundin wäre durch
diese Freundschaft ranghoch, hätte also einen von dieser Beziehung abhängigen hohen
Rang. Eine solche Erklärung stünde im deutlichen Gegensatz zu unserem Eindruck,
dass nur bestimmte Typen von Weibchen es schafften, freundschaftliche Beziehungen
zum Alpha-Weibchen zu halten.
Zur Klärung führten wir umfangreiche Separations- und Introduktionsversuche durch
(Unterkapitel 14.3). Erstes Ergebnis unserer damaligen Experimente war, dass alle
Tiere nach vierzehntägiger Separation ihre alte Rangposition in der Gruppe
zurückerhielten. Dies galt für ranghohe, rangmittlere und rangniedrige Individuen.
Darüber hinaus änderten die Separationen auch nichts an den Beziehungen der übrigen
Gruppenmitglieder, selbst als wir Alpha-Männchen und Alpha-Weibchen gemeinsam
separierten. Die einmal etablierte Rangordnung schien nahezu nicht manipulierbar zu
sein. Ziel des weiteren Vorgehens war es daher, diese festgefügte Ordnung
experimentell zu ändern.
Um den rangniedrigen Individuen eine bessere Startposition zu geben, separierten
wir die niederrangigsten Clane (Mütter gemeinsam mit ihren in der Gruppe
geborenen Kindern) und introduzierten sukzessive in die somit neu etablierte
Gruppe die anderen Clane entgegengesetzt zur bisherigen, gut etablierten
Rangordnung.
Es war nun bemerkenswert, dass die früher rangniedrigen Individuen in der Lage
waren, alle Verhaltensweisen zu zeigen und im richtigen Kontext anzuwenden, die
üblicherweise nur an hochrangigen Individuen zu beobachten sind. Diese Fähigkeit
besaßen sie offensichtlich unabhängig von ihrer früheren Rangposition in der Gruppe.
Im Gegensatz zu den bereits erwähnten Resultaten der vorher durchgeführten
Reintroduktionsexperimente war es nun jedoch auffällig, dass sich die Individuen bei
Introduktion in die Gruppe, in der die früher rangniedrigsten ranghoch waren,
bezüglich ihres Verhaltens unterscheiden ließen.
„Rangniedrige“ Individuen erhielten eine Position unter den künstlich ranghohen Das
gleiche galt für „rangmittlere“ Individuen, die ebenfalls nicht in der Lage waren, die
künstlich Ranghohen zu dominieren. Aber im Gegensatz zu den Rangniedrigen griffen
die Rangmittleren die Individuen an, die einen Experimentalschritt früher introduziert
wurden, und sicherten sich hierdurch diesen gegenüber ihre alte Rangposition. Wir
nannten dieses Verhalten „Rangsicherungsverhalten“.
Ganz im Gegensatz zu den rangniedrigen und rangmittleren Tieren hatten ranghohe
Individuen keine Probleme, ihre hohe Position wieder zu erhalten. Es war nun aber
besonders bemerkenswert, dass in der gleichen Zeit, in der die hochrangigen Individuen
ihren Rang etablierten, die rangmittleren die künstlich ranghohen attackierten. Wir
nannten dieses Verhalten „Rangverbesserungsverhalten“.
Auch ein Kontrollexperiment erbrachte die gleichen Resultate. Wir hatten also
gezeigt, dass die endliche Position jedes Individuums von seinem Verhalten in neuen
sozialen Situationen abhängig war. Differenzieren wir zwischen „ranghoch“,
„rangmittel“ und „rangniedrig“, dann war der erreichte Rang, den wir als natürlichen
Rang bezeichnet haben ([242]), demnach eine Eigenschaft des Individuums.
Hochrangige Individuen waren in der Lage, ihre hohe Position zu halten, mittelrangige
zeigten „Rangsicherungs“ und „Rangverbesserungsverhalten“, niedrigrangige
waren nur in der Lage, eine etablierte soziale Position zu halten, zeigten aber
keine sozialen Strategien, ihre Positionen zu verbessern oder zu verteidigen.
Wir waren jedoch nicht in der Lage, zu entscheiden, ob diese Eigenschaft auf
früheren sozialen Erfahrungen beruhte oder aber eine angeborene Disposition
war.
Wir hatten nämlich keine Aussage darüber gemacht, ob für die Eigenschaft Rang nun
Erziehungsprozesse in der Gruppe verantwortlich sind, oder aber noch zu
bestimmende, vielleicht auch genetische Effekte. Erziehungsprozesse in der Gruppe
könnten ja theoretisch das zukünftige Verhalten und damit das zukünftige
Leben soweit beeinflussen, dass der Geburtsrang sich in der Zukunft verewigen
würde. Wir mussten also Erziehungsprozesse durch Mütter und Geschwister
ausschließen.
Für Experimente konnten wir statt der Adulten auch in der Gruppe geborene
Jungtiere nutzen. Wie bereits oben erwähnt, finden wir nämlich bei Makaken generell
die Situation, dass in der Gruppe geborene Jungtiere den relativen Rang der Mutter
übernehmen, d. h. sie erben sozusagen den Rang der Mutter, unabhängig vom
Zeitpunkt des Geborenwerdens. Jüngere Kinder ranghoher Mütter sind demnach
ranghöher als weitaus ältere rangniedrigerer Mütter.
Am sinnvollsten erschien es uns, Tiere zu wählen, die postnatal ohne Einflüsse von
Müttern und Geschwistern aufgezogen wurden. Wir separierten daher 1982 und 1983
alle in der Gruppe geborenen Jungtiere gleich nach der Geburt und zogen sie in einer
„peer-group“ auf, einer Spielgruppe mehr oder weniger gleichalter Kinder, wie sie als
Experimentalgruppen auch aus amerikanischen Primatenzentren bekannt sind
(Unterkapitel 14.6).
Um Deprivationssyndrome zu verhindern, wurde jedes Tier in den ersten vier
Lebenswochen individuell von nur jeweils einem meiner Mitarbeiter betreut.
Nach dieser Periode überführten wir die Jungen in einen Innenkäfig unserer
Handaufzuchtanlage, die aus einem größerem Spielkäfig und einem kleineren
Innenkäfig bestand. Hier hatten die vier Wochen alten Jungtiere auch erstmals
Gitterkontakt zu Artgenossen. Nach einer Woche in diesem Innenkäfig, also
sobald das jeweilige Jungtier fünf Wochen alt war, wurde die Tier geöffnet
und das Jungtier zu den anderen versuchsplanbedingt älteren Jungtieren
gelassen.
Generell wurden alle Jungtiere nach dem gleichen Muster aufgezogen wenngleich die
Aufzuchtsanlage erheblich modifiziert werden musste. Die ursprüngliche Anlage
hat sich nämlich nicht lange bewährt. Als die Tiere älter wurden, warteten
sie geradezu auf die Chance, den Käfig zu verlassen, und die Umgebung,
mein Arbeitszimmer, zu erkunden (und zu zerstören). So mussten wir die
Versuchsapparatur ändern, durch einen zusätzlichen Käfig und schließlich durch weitere
Absperrmöglichkeiten. Das Endergebnis war dann eine Apparatur aus zwei miteinander
verbundenen Käfigen und einem Futterturm zum Separieren und „handling“der
Tiere.10
Bei normalen Handaufzuchtsmethoden werden die jungen Makaken gleich
nach Geburt separiert, für eine gewisse Zeit separat gehalten und zu einem
festen Zeitpunkt gemeinsam mit anderen Individuen gleicher Vorgeschichte in
einen „peer-group“Käfig überführt. Wir hingegen introduzierten einen Affen
nach dem anderen, sobald das jeweilige Individuum fünf Wochen alt war.
Insgesamt bildeten wir zwei „peer-groups“, eine mit den 1982, die andere mit den
1983 geborenen Jungtieren. Die erste bestand aus acht, die zweite aus zwölf
Individuen.
Zum „handling“ der Tiere sei bemerkt, dass wir bis zu dem Tag, an
dem das jeweils jüngste Individuum sechs Monate alt wurde, allen Tieren
fünfmal pro Tag eine Flaschenmahlzeit gaben. Ältere Jungtiere erhielten
täglich einmal die Flasche. Hierdurch blieben die Tiere vertraut mit der
„handling“-Prozedur.11
Nach den Befunden an anderen „peer“-Gruppen mussten wir als mögliche Parameter
der sozialen Entwicklung das Geschlecht, das Alter und das Gewicht Tiere
berücksichtigen.
Zusätzlich kam bei unseren Gruppen, und insofern unterschieden sich unsere
Experimentalgruppen von allen anderen vergleichbaren, als möglicher Parameter auch
noch der Rang der jeweiligen Mutter in der Ausgangsgruppe, der in unserer Kolonie
umfänglich protokolliert und getestet wurde, in Frage.
Zum Einfluss des Geschlechtes fanden wir, dass es in beiden Geschlechtern zwar
schwerere und leichtere Individuen gibt, Männchen jedoch durchgängig im Mittel
schwerer sind als Weibchen. Ansonsten kam dem Geschlecht keine große Bedeutung zu,
wie nach den Literaturberichten - Männchen sollen ranghöher sein als Weibchen - zu
erwarten. Männchen fügten sich in die Hierarchie ein, dominierten also jüngere und
waren subordinant gegenüber älteren Individuen.
Damit wäre auch schon der entscheidende Paramter für die erste Lebensphase
genannt, hier kommt offensichtlich dem Alter höchste Bedeutung zu.
So fanden wir, dass die Jungtiere eine altersabhängige Rangordnung ausbildeten,
unabhängig von Geschlecht und Gewicht. Ohne jede Ausnahme waren ältere Tiere
höherrangig als jüngere.
Es war zudem auffällig, dass alle Individuen enge Kontakte zu Tieren des gleichen
Altersbereiches zeigten, unabhängig vom Geschlecht.
Beachten wir, dass jüngere Tiere später in ihre „peer-group“ kamen als ältere, dann
waren diese Resultate die nämlichen, die wir zehn Jahre vorher erhalten hatten, als wir
unsere Zuchtgruppe aus Wildfängen bildeten. Fanden wir doch damals, dass die Tiere
entsprechend dem Introduktionszeitpunkt eine Hierarchie ausbildeten, dass
also früher introduzierte Tiere anfänglich einen höheren Rang erhielten als
später introduzierte (vgl. Unterkapitel 14.2). Zudem fanden wir, dass Tiere
benachbarter Ränge enge Beziehungen zueinander aufnahmen. Vergleichen wir diese
alten Resultate mit den neuen, dann schienen die Mechanismen die selben zu
sein.
In der bereits mehrfach erwähnten Ausgangsgruppe aus adulten Individuen, mit denen
wir unsere Untersuchungen begannen, fanden wir, dass die Tiere sich unterschiedlich
verhielten, als sie in die Gruppe introduziert wurden. In Abhängigkeit von ihrem
Verhalten verbesserten einige ihre soziale Position, andere waren hierzu nicht in der
Lage. Offen war nun die Frage. ob identische oder vergleichbare Prozesse auch in den
„peer-groups“ ablaufen würden. Unsere Frage war also, ob sich diese Rangordnung
verändern würde. Wir müssen dabei beachten. dass es viel leichter ist, einen
einmal erhaltenen Rang zu halten, als ihn zu verbessern, wofür vornehmlich das
rangstabilisierende Interaktionssystem, alle Tiere halten bevorzugt Kontakt zum
jeweils Rangnächsten, verantwortlich ist. Analysen der beobachteten Änderungen([296],
[284]) ergaben folgende Tendenzen: Individuen, die ihren Rang verbessern, haben
ranghohe Mütter, diejenigen, die ihren Rang verschlechtern, rangniedrige
Mütter in der Geburtsgruppe. Ausnahmen von diesem „Trend“ konnten nicht
beobachtet werden. Nach der Fusion beider „peer“- Gruppen war dann die
endliche Rangordnung nahezu identisch mit derjenigen, die zu erwarten gewesen
wäre, hätten wir auf die Separation dieser Individuen aus ihrer Geburtsgruppe
verzichtet.
Eine naheliegende Frage war nun, wodurch man ranghoch bzw. rangniedrig werden
würde, auch hier sind Aussagen möglich. Das „Rangniedrigwerden“ verlief nach einer
einfachen Regel, so verloren Tiere ihren Rang, die häufig und - nach dem sozialen
Kontext - unnötig andere bedrohten. Sie bedrohten nämlich Individuen, die
rangniedriger waren, submissives Verhalten zeigten und die durch ihr Verhalten
in einer normalen Gruppensituation eigentlich agonistisches Verhalten der
anderen verhindern würden. Hier zeigten rangniedrige offensichtlich falsche
Antworten.
Weitaus schwieriger erschien der Weg, „ranghöher“ zu werden. Auffällig war, dass ein
seinen Rang verbesserndes Individuum scheinbar nichts unternahm, um seinen Rang
zu verbessern, in Wirklichkeit aber verfolgt es gleichzeitig vier verschiedene
Strategien:
(1) Vermeidung jedes gegen sich selbst gerichteten agonistischen Verhaltens. Dies
erreichte das Individuum durch submissives Verhalten gegenüber Ranghöheren, durch
das Negieren des Bedrohtwerdens bzw. durch die Aufnahme besonders enger sozialer
Kontakte zu dem ranghöheren Tier, das das Individuum eben gerade bedroht
hat.
(2) Aufwertung des eigenen „Wertes“ durch agonistische Auseinandersetzungen
beendendes Verhalten. Dies erreichte das Individuum durch scheinbar planlos und
unbeteiligtes Herumlaufen im Käfig, wobei es jedoch tatsächlich mehrfach sich vor ein
gerade bedrohtes Tier setzte, wodurch das gegen dieses gerichtete agonistische
Verhalten beendet wurde. Das bedrohte Individuum suchte danach die Nähe des
„Schlichters“ und wertete diesen auf. Soweit der Aggressor rangniedriger war als der
„Schlichter“, konnten wir auch direkte Interventionen beobachten, war er
ranghöher, blieb der „Schlichter“ scheinbar völlig unbeteiligt und passiv. Er
erreichte aber, dass das angegriffene Tier - zumindest mittelfristig - ihm selbst
unterlegen wurde. Erkennt dieses nämlich den „Schlichter“ nicht als ranghöher
an, reichte eine kurze Koalition mit dem angreifenden Tier, um die neuen
Rangverhältnisse zu stabilisieren. Der „Schlichter“ war also ohne Aufwand ranghöher
geworden.
(3) Bildung von „Freundschaften“ mit ranghöheren Individuen. Dies erreichte das
seinen Rang letztendlich verbessernde Individuum durch das aktive Aufsuchen Putzen
und Umarmen Ranghöherer, unabhängig von deren aktueller Antwort; d. h., auch
wenn der Ranghöhere die Nähe mied bzw. sogar dem Individuum drohte, bliebt er Ziel
für Sozialkontakte.
(4) Befriedung der Unterlegenen. Dies erreichte das Individuum durch ein scheinbar
paradoxes Verhalten: Unterlegene Individuen wurden verfolgt und mehrfach gebissen,
bis sie wirklich jegliche Gegenwehr vermieden, dann jedoch wurden sie Ziel engster
Sozialkontakte, geputzt und umarmt wie Ranghöhere. Die Kombination aller
vier Strategien gleichzeitig zeigten nur Individuen. die ranghöher wurden.
Einzelne Strategien konnten jedoch von jedem Gruppenmitglied angewandt
werden.
Offensichtlich kommt es nicht auf die Quantität sondern auf die Qualität des
agonistischen Verhaltens an. Das Individuum muss wohl das agonistische
Verhalten zur richtigen Zeit anwenden. Viele aggressive Handlungen erlauben eher
Negativ-Prognosen für die Zukunft.
Wir hatten also gezeigt, dass offensichtlich dem Rang der Mutter hohe Bedeutung
zukommt, auch wenn das Jungtier die Mutter postnatal überhaupt nicht erfahren
konnte, hatten wir doch ihren Einfluss und denjenigen weiterer Geschwister
ausgeschlossen.
Über die Bedeutung des Rangplatzes des Vaters ist damit keine Aussage gemacht. Da
in unserer Kolonie nur ein Männchen als Vater in Frage kam, konnten wir dessen
Einfluss vernachlässigen.
Über den Einfluss des Vaters gaben jedoch zusätzliche Kontrollbeobachtungen an einer
zweiten Makakenspecies, dem Rhesusaffen, Auskunft, die ich gemeinsam mit Elke
Harigel 1984 an dem ehemaligen Harlow-Institut in Madison, Wisconsin, durchführen
durfte (Unterkapitel 14.7).
Männchen und Weibchen scheinen demnach etwas zu vererben, was für den späteren
Rang des Individuums von Bedeutung ist.
Der Rang ist dabei nur ein Maß für die Fähigkeit des Individuums, adäquat seinem
Sozialpartner zu antworten. Diese Fähigkeit hatten wir soziale Intelligenz
genannt.
Zudem wagten wir einen neuen, weiteren Forschungsschritt. Wir gingen davon aus,
dass dem Rang der Mutter bei gleich nach Geburt separierten Kindern entscheidende
Bedeutung zukommt, dass also postnatale Einflüsse vernachlässigt werden können.
Insofern würde es notwendig sein, Fragen nach möglichen pränatalen Einflüssen
nachzugehen. Hierzu teilten wir unsere Zuchtgruppe in drei neue Gruppen auf, in eine
mit nur ranghohen, eine mit nur rangmittleren und eine mit nur rangniedrigen
Individuen. Wir konnten davon ausgehen, dass sich in jeder dieser drei Gruppen eine
lineare Rangordnung etablieren wird und dass es in jeder Gruppe ranghohe und
rangniedrige Individuen geben wird. In der Gruppe der Ranghohen wird es also Tiere
geben, die definitionsgemäß zwar ranghoch sind, dennoch aber erfahren, rangniedrig
zu sein. In der Gruppe der Rangniedrigen hingegen werden entsprechend
Rangniedrige ranghoch sein. Hierdurch konnten wir die pränatalen Einflüsse
manipulieren. Den Einfluss gerade der pränatalen Phase sollte abschätzbar
sein.
Gehen wir davon aus, dass dem Vater wirklich Bedeutung zukommt, dann müssten
sich durch diese Versuchsanordnung zudem Einflüsse von Vater und Mutter addieren.
Auf die Ergebnisse und die erheblichen Schwierigkeiten unserer Bemühungen werde ich
in Unterkapitel 14.9 eingehen.
Zudem gebe ich Informationen zu unserem Experiment zum Verwandtenerkennen
(Unterkapitel 14.10). Schließlich stelle ich unsere Befunden über Hormone und
Verhalten (Unterkapitel 14.11) und über Lernexperimente (Unterkapitel 14.12) vor.
Sowohl bei der Hormonsekretion (Hypophysen-Nebennierenrinden-Achse) als auch
bei den Lernversuchen können wir Ranghohe und Rangniedrige eindeutig
diskrimieren.
Gemeinsam mit meiner Mitarbeiterin Annette Klaiber-Schuh wertete ich 1995 die
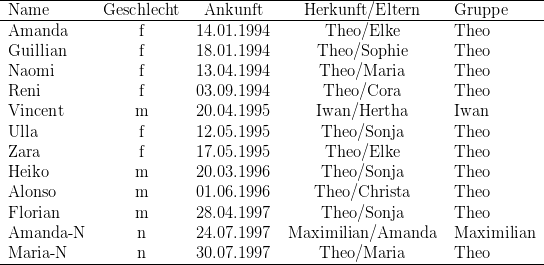
Reproduktionsdaten unserer Javanermakaken aus ([267]). Insgesamt wurden bis zum
Zeitpunkt der Auswertung 196 Kinder (nur Einlinge) geboren. 84 % der Neugeborenen
überlebten die erste Lebenswoche (bei den importierten Weibchen sogar 90 %), von
den Überlebenden erreichten dann 84 % zumindest das erste Lebensjahr. Von ihren
Müttern nicht angenommene und daher handaufgezogene Individuen wurden als tote,
experimentell handaufgezogene (ihren Müttern abgenommene) Kinder als lebende
Individuen gezählt. Erstgebärende von der Mutter aufgezogene Weibchen (n = 15)
waren ähnlich erfolgreich wie mehrfachgebärende, 66 % der Kinder überlebten. Bei den
handaufgezogene erstgebärenden Weibchen dagegen überlebten nur 20 %.
Die hohe Todesrate der Kinder von handaufgezogenen Müttern war nicht durch
mangelndes mütterliches Verhalten bedingt, vielmehr starben die Jungtiere durch
„kidnapping“. Die anderen handaufgezogenen Weibchen respektierten anfänglich nicht
den Jungenbesitz, vielmehr raubten sie die Jungen und pflegten sie fürsorglich zu
Tode.
Das mittlere Alter der 27 erstgebärenden Weibchen war 5 Jahre 5 Monate (± 2
Monate). Das jüngste Weibchen war am Tag der Geburt 4 Jahre und fünf Monate
alt.
Der Geburtenabstand hängt deutlich von dem Aufzuchtserfolg ab, er betrug 473 ± 14
Tage bei Kinder tragenden Weibchen (n = 120) und 384 ± 27 Tage bei Weibchen, die
das Kind verloren hatten (n = 62). Eine Saisonalität war bei den Geburten in der
Kolonie nicht auffällig.
Das Geschlechterverhältnis Männchen zu Weibchen betrug 1 : 0,89 bei allen
Jungtieren.
Da unsere Ergebnisse auf dem Verhalten von Makaken-Persönlichkeiten beruhen, gebe
ich in den folgenden Unterkapiteln die Beiträge dieser Individuen an der Entstehung
unseres Wissenstandes an. Dabei verzichte ich weitgehend auf graphische
Darstellungen, diese sind in den zitierten Publikationen zugänglich.
Zu Beginn unserer Langzeitstudie zum Sozialverhalten von Macaca fascicularis entschlossen
wir uns zum sukzessiven Aufbau der Gruppe. Hierdurch konnten wir die Interaktionen
eines jeden Individuums und seine Strategien, eine höhere soziale Position zu erreichen,
aufzeigen.12
Es schien uns sinnvoll, um die soziale Stellung rangniedriger Weibchen unabhängig von
dem Beisein des Männchens testen und eventuelle Strategien der Weibchen,
eine höhere soziale Position zu erlangen, erkennen zu können, im Verlauf der
Introduktionen einige Weibchen zu separieren, bzw. in eine nur aus Weibchen
bestehende Gruppe zu introduzieren.
Die Beschränkung auf nur ein adultes Männchen erlaubte bei den weiteren
Untersuchungen, den Einfluß des Vaters auf verwandtschaftliche Beziehungen vorerst
vernachlässigen zu können, waren doch alle in der Gruppe geborenen Jungtiere
Halbgeschwister zueinander.
Eine Situation wie in unserem Labor, nämlich dass Weibchen sukzessive in eine
bestehende Gruppe introduziert werden, wird es im Freiland kaum geben, doch sah ich
gerade durch eine solche Versuchsanordnung die Möglichkeit, die Komplexität des
Verhaltens und die Bedingtheiten eines jeden Individuums besser abschätzen zu
können.
14 der 16 an dieser Untersuchung teilnehmenden Individuen waren Wildfänge, zwei vor Untersuchungsbeginn in Kassel geboren. Vier der Wildfänge erhielten wir von den Behring-Werken in Marburg, wo die Tiere bereits quarantänisiert und einige Monate gehalten wurden.13
Die Javaner-Affen beobachtete einer von zwei - den Makaken vertrauten - Beobachtern täglich im Innenraum und protokollierte ihre Interaktionen. Nach Introduktionen wurden die neu introduzierten Tiere und die Gruppe darüber hinaus mindestens eine Stunde direkt beobachtet, dabei saß der/die Beobachter/in direkt vor den Käfigen; zusätzlich wurden von einigen Introduktionen Videoaufzeichnungen angefertigt.
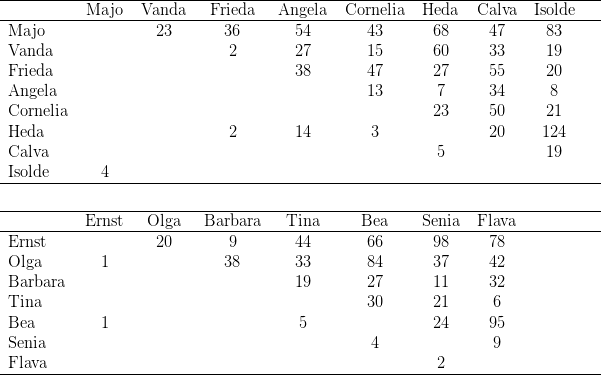
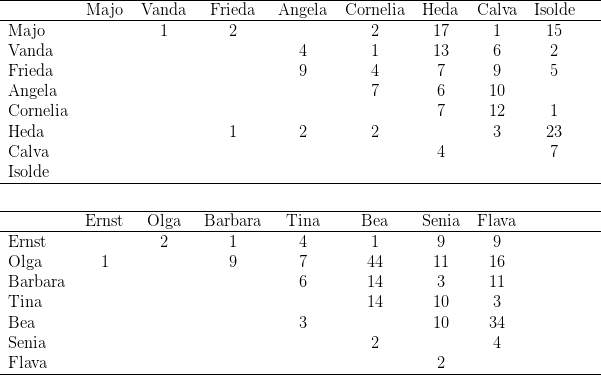
Zur Dominanzbestimmung separierten wir jeweils zwei Tiere in einen Käfig und legten auf den Boden des Käfigs ein Stück Ei oder eine Nuss. Protokolliert wurde, welches der Tiere sich das Futter holte und welche Interaktionen anschließend erfolgten. Diese Versuche führten wir stets dreimal hintereinander durch, hierbei jeweils alle möglichen Zweierpaarungen testend.14
Bei der Introduktion adulter weiblicher Macaca fascicularis in die Nucleusgruppe war
stets zu beobachten, dass das Männchen Nikita das neuintroduzierte Weibchen (1)
verfolgte, bis dieses (2) präsentierte; daraufhin inspizierte er das Weibchen, (3) stieg
auf und (4) biss es in den Rücken. Diese vier Stadien verliefen meist in der gleichen
Reihenfolge, nur bei Jungfrau folgte das vierte Stadium „Beißen“ gleich nach dem
ersten Stadium „Verfolgen“, was wohl hinreichend durch das Nichtpräsentieren
Jungfraus erklärt werden kann. Dieses abweichende Verhalten könnte eventuell durch
Jungfraus Alter bedingt sein.
Vergleichen wir diese Beobachtungen mit denen Kummers ([117]) an
BlutbrustpavianenTheropithecus gelada, so gilt für die Introduktionen bei Macaca
fascicularis im Prinzip auch die für Theropithecus gefundene „Regel 1“: „A
dyad of adult geladas goes through four stages. Stage 1 is fighting, stage 2
presenting, stage 3 mounting and stage 4 grooming“ ([117], Seite 130). Agonistische
Interaktionen gingen jeweils einseitig von Nikita aus, auch wehrte sich mit
Ausnahme von Barbara keines der Weibchen. Hierdurch bedingt war Stadium 1
stets „Verfolgen“. Stadium 2 und Stadium 3 sind identisch mit Kummers
Stadien. Als neues Stadium kommt bei Macaca fascicularis „Beißen in den
Rücken“ hinzu. Kummers Stadium 4 „grooming“ war bei Macaca fascicularis
ebenfalls vorhanden.
Die Javanermakaken erreichten dieses 5. Stadium der Integration (Soziale Körperpflege)
erst viel später, was wiederum auf die Gruppensituation zurückgeführt werden
kann.15
Diese dyadische Interaktion Nikita/neues Weibchen wurde zum gleichen Zeitpunkt
überlagert durch die dyadische Interaktion Vanda/neues Weibchen, welche
sich vor allem in dem Beißen des Alpha-Weibchens äußerte. Der Biss Vandas
konnte vor oder nach dem Beißen des Männchens stattfinden, er ereignete
sich stets dann, wenn das neuintroduzierte Weibchen auf der Flucht vor dem
Männchen sich dem ruhig, scheinbar teilnahmslos sitzenden Alpha-Weibchen
näherte.
Die gleichen Verhaltenssequenzen galten auch für Introduktionen in die
Weibchen-Gruppe bzw. für das Zusammenlassen ranghöherer mit einem rangniedrigeren
Weibchen. Hier konnte gleichfalls neben den Stadien 1 und 2 auch die Stadien 3 - 5
beobachtet werden, nur mit stärkerer zeitlicher Verzögerung.
Ein auffallender Unterschied zwischen Introduktionen in die Nikita- und in
die Weibchen-Gruppe war jedoch, dass in der Nur-Weibchen-Gruppe alle
Weibchen an agonistischen Interaktionen beteiligt waren, während sich in der
Ein-Männchen-Gruppe agonistische Interaktionen auf Nikita und Vanda beschränkten.
Durch die Anwesenheit des Männchens werden somit Interaktionen zwischen den
Weibchen reguliert, dem Männchen kommt also eine Kontrollfunktion bei
Gruppenauseinandersetzungen zu.
Reintroduktionen verlaufen friedlicher. Von den fünf Stadien entfallen meist Stadium
1 (Verfolgen) und Stadium 4 (in den Rücken beißen), manchmal auch Stadium 3
(Besteigen). Für den Verlauf der Reintroduktionen kommt dem Verhalten der
reintroduzierten Weibchen erhebliche Bedeutung zu. Unabhängig hiervon finden wir
jedoch drei verschiedene Phasen der Interaktionen. So nehmen reintroduzierte
Weibchen nacheinander aktiv oder passiv Kontakt zu den ranghohen Tieren der
Gruppe (1. Phase), dann zu den anderen Gruppenmitgliedern (2. Phase) auf, um dann
anschließend ihren Rang zu sichern (3. Phase).
Um die unterschiedlichen Reaktionen der Individuen zu belegen, bzw. die
Rangänderungen nachvollziehbar zu machen, beschreiben wir hier alle Versuchsabschnitte
chronologisch. Bemerkt werden soll noch, dass zum Zeitpunkt der jeweiligen
Introduktion kein Weibchen oestrisch war, wobei grundstzlich betont werden
muss, dass ein Oestrus grundsätzlich keinen Einfluss auf die Rangordnung
hat.
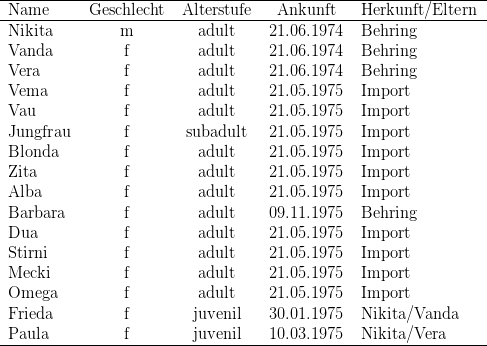
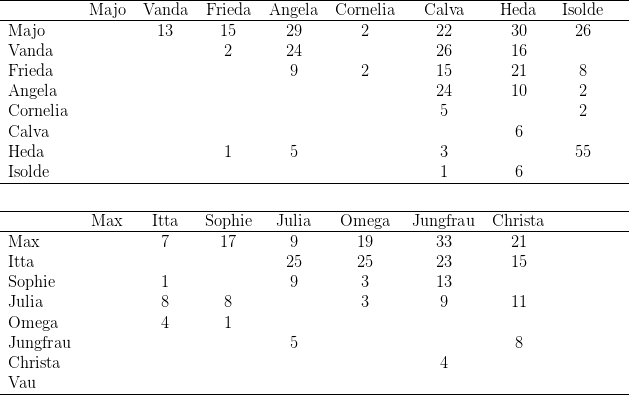
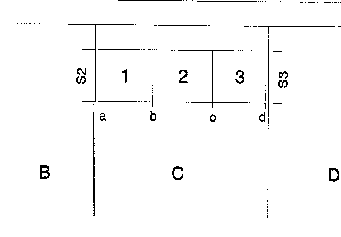
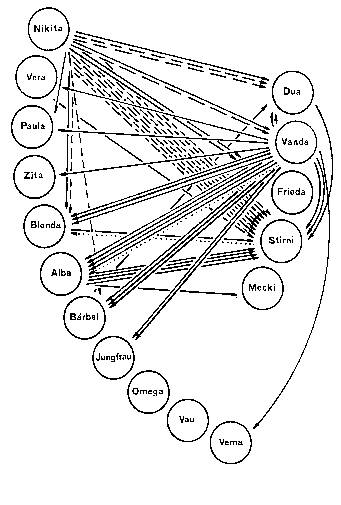
Kerngruppe Die Kerngruppe unserer Macaca fascicularis - Kolonie bestand aus dem Männchen Nikita und den Weibchen Vanda und Vera. Wir erhielten die drei Tiere am 21. 06. 1974.
Nikita und Vanda wurden bereits vorher gemeinsam gehalten. Vom ersten Tag an dominierte Vanda deutlich über Vera, was sich besonders bei der Nahrungsaufnahme zeigte. Auch war Vanda an agonistischen Interaktionen kaum beteiligt, sie saß vielmehr in der Regel ruhig auf einem im Hintergrund des Käfigs angebrachten Sitzbrett. Vera hingegen wurde bei vermeintlicher Bedrohung von Nikita gebissen und stürzte daraufhin an die Vorderfront des Käfigs, um der „Störung“ (Beobachter, Tierpfleger) zu drohen. An diesem Zustand änderte sich auch nichts, als im Januar bzw. März 1975 beide Weibchen ein Junges bekamen.
Introduktion von Vema
Die am 21.05.1975 importierten Macaca fascicularis-Weibchen wurden vom ersten Tag
ihrer Ankunft in Kassel an gemeinsam mit Nikita, Vanda und Vera in einem Raum,
jedoch in getrennten Käfigen gehalten. Am 22. Juni 1975, also knapp vier
Wochen nach Import, introduzierten wir das erste „neue“ Weibchen in die
Kerngruppe.
Nach kurzer Verfolgungsjagd und olfaktorischer Kontrolle der Anogenitalregion
(Inspizieren ([2])) stieg das Männchen auf und biß Vema in den Rücken. Anschließend
biss Vanda ebenfalls mehrfach das neuintroduzierte Weibchen. Vema blieb hierbei mit
zur Wand gewandtem Gesicht sitzen, bzw. zeigte das Pressen, d. h. sie blieb mit
angezogenen Gliedmassen, steif, flach bäuchlings am Boden liegen. Verletzungen
konnten trotz vieler Bisse nicht festgestellt werden. Hier, wie auch bei den folgenden
Introduktionen, packten sowohl Nikita als auch Vanda das Fell nur mit den Incisivi
und zogen. Verletzungen resultierten dabei wohl eher aus einem Reißen der Haut als
von einem Durchbiss.
Entsprechend der Introduktionsfolge nahm Vema jetzt den dritten, letzten Rang
innerhalb der Weibchen ein. Rangbestimmungen erfolgten im weiteren durch
regelmässige Futterkonkurrenztests.
Separierung von Vera und Paula Am 02.07.1975 separierten wir das Beta-Weibchen, Vera, gemeinsam mit seiner Tochter Paula. An der Gruppenstruktur änderte sich nichts. Vanda blieb erwartungsgemäß das Alpha-Weibchen.
Sukzessive Introduktion der vorher gemeinsam gehaltenen Weibchen Vau,
Jungfrau, Blonda und Zita
Vom 07. - 11. Juli 1975 introduzierten wir sukzessive die restlichen vier im
benachbarten Käfig sitzenden Weibchen. Vor der Introduktion bestimmten wir ihre
Rangfolge und ließen sie dann in umgekehrter Rangfolge zu. Alle vier Weibchen
wurden zur Introduktion jeweils in einen Umsetzkäfig mit Kantenlänge 40 cm gesperrt,
der in den Käfig der Kerngruppe 30 Minuten vor Freilassen des Weibchens gestellt
wurde.
Allen vier Introduktionen war gemeinsam, (1) dass das Männchen während dieser
halben Stunde nervös im Käfig herumlief, dabei zum einen jedes seiner Weibchen
bedrohte,16
zum anderen den Umsetzkäfig optisch und olfaktorisch
untersuchte,17
(2) dass das Männchen nach dem Freilassen die Weibchen verfolgte und nach ihnen
griff, um die Anogenitalregion olfaktorisch zu prüfen, aufzusteigen und die Weibchen in
den Rücken zu beißen, (3) dass die Weibchen stets auch von dem Alpha-Weibchen,
Vanda, gebissen wurden.
Unterschiedlich war hier nur die individuelle Reaktion der Weibchen und damit auch
der Ablauf der Introduktionen. Das am 07.07.1975 erstintroduzierte Weibchen, Vau,
ließ sich widerstandslos inspizieren, beißen und besteigen (Imponierbegatten ([2])) und
blieb, wie zuvor schon Vema, in Demutsstellung (Stummes Pressen([2])) sitzen. Das
am 08.07.1975 nächstintroduzierte Weibchen, Jungfrau, dagegen ließ das
Männchen nicht aufsteigen, vielmehr floh sie ständig vor Nikita und Vanda durch
den Käfig, was zu ständiger Verfolgung und zu unzähligen Bissen sowohl
durch Nikita als auch durch Vanda führte. Das erste Aufsteigen konnte erst
knapp ein Jahr nach der Introduktion beobachtet werden. Die am 10.07.1975
introduzierte Blonda, das dritte Weibchen, hingegen blieb wiederum sitzen und floh
nicht. 13 Sekunden nach Freilassen wurde sie bereits von Nikita und Vanda
gebissen, worauf sie sofort präsentierte und im folgenden die Nähe Vandas
suchte.
Bereits am nächsten Tag wurde der unterschiedliche Integrationsgrad der letzten
beiden Weibchen deutlich, Blonda hielt sich stets neben Vanda auf, Jungfrau dagegen
wurde ständig von Nikita verfolgt; sie versuchte, ihm auszuweichen, stand somit dem
wohl angestrebten Ziel „Zusammenhalten der Gruppe“ entgegen und rief dadurch
stets neue agonistische Interaktionen hervor. Das am 11.07.1975 introduzierte
Weibchen Zita wiederum floh anfänglich auch vor Nikita, bis sie vier Minuten
nach Freilassen von Vanda gepackt und gebissen wurde. Hierauf blieb sie in
Demutstellung sitzen und ließ sich widerstandslos von Nikita inspizieren.
Das erste Aufsteigen fand jedoch erst sechzehn Minuten nach Introduktion
statt.
Reintroduktion von Vera und Paula Bei der am 15.08.1975 erfolgten Reintroduktion von Vera und Paula fanden keine agonistischen Interaktionen statt, Vera nahm ohne Rangstreitigkeiten wieder ihre alte Position nach Vanda in der Weibchenhierarchie ein. Nikita beschränkte sich auf bloßes Inspizieren des bei Annäherung sofort präsentierenden Weibchens. Für die zweite Position innerhalb der Weibchen kann die physische Kraft als Grund ausgeschlossen werden, verlor doch Vera durch eine Operation18 während der Separierung die Incisivi und Canini des Unterkiefers.
Introduktion von Alba Am 17.09.1975 wurde dann Alba, ein ranghohes Weibchen der Weibchen-Gruppe, introduziert. Der Verlauf des Geschehens war der Nämliche. Nach kurzer Verfolgung durch Nikita wurde Alba inspiziert, gebissen und bestiegen; nach sechsmaligem Aufsteigen ließ Nikita von Alba ab, worauf sich Vanda sofort auf sie stürzte, Alba in den Rücken biss und anschließend das fliehende Tier verfolgte.
Introduktion von Barbara
Da wir nicht ausschließen konnten, dass die gemeinsame Haltung aller Makaken in
einem Raum und das dadurch bedingte Sichkennen verantwortlich für die beobachteten
Interaktionen war, introduzierten wir als nächstes ein neu von den Behring-Werken
übernommenes Weibchen, das bis zum Moment der Introduktion keinen Kontakt zu
unserer Makaken-Kolonie gehabt hatte.
Dieses Weibchen, Barbara, wurde am 10.11.1975 die ersten 20 Minuten
ausschließlich mit Nikita zusammengesperrt. Dieser näherte sich, wie bei allen
anderen Introduktionen, dem Weibchen, wurde aber von Barbara angegriffen
und wich vor ihr. Ein Vorgang, der sich in den ersten zwei Minuten nach
Introduktion dreimal wiederholte. Darauf folgte eine 12-minütige Phase von
Inspektionen und Aufsteigen (Angst: Imponierbegatten), wobei jedoch erst 13
Minuten nach Introduktion der Rückenbiss erfolgte. Nach einer 5-minütigen
Phase geringer sozialer Aktivität schloss sich eine kurze Phase fortwährenden
Beißens an, die zum Pressen des Weibchens führte und wohl endgültig die
besondere Stellung des Männchens festigte. Hierauf ließen wir die übrige Gruppe
zu und konnten bereits nach 10 Minuten den Zubiss des Alpha-Weibchens
beobachten. In den ersten 35 Minuten nach Vergesellschaftung mit der übrigen
Gruppe zeigte Barbara das Pressen und ließ sich widerstandslos von Nikita
inspizieren. Im Folgenden konnten weitere agonistisches Verhalten Aktivitäten
Nikita/Barbara beobachtet werden. Daneben war auffällig, dass Vanda, Vera und Zita
versuchten, sich dem neuen Weibchen zu nähern, doch wurden sie stets von Nikita
verjagt.
Introduktion von Vau in die Weibchengruppe Am 02.02.1976, die Gruppenstrukturen hatten sich gefestigt, überführten wir das rangniedrigste Weibchen der Nikita-Gruppe, Vau, in die nur aus Weibchen bestehende Gruppe. Hier wurde sie sofort von den beiden ranghöchsten Weibchen, Dua und Stirni, angegriffen und in den Rücken gebissen, Vau blieb hierbei in Demutsstellung sitzen. Am nächsten Tag konnten auch agonistische Aktivitäten des Gamma-Weibchens, Mecki, beobachtet werden, das rangtiefste Weibchen, Omega, hingegen griff Vau nicht an, agonistische Aktivitäten Vaus waren ebenfalls nicht zu beobachten, es war jedoch auffällig, dass sie den angreifenden Weibchen präsentierte.
Introduktion von Vanda in die Weibchengruppe
Am 12.02.1976 introduzierten wir das Alpha-Weibchen, Vanda, in die Weibchen-Gruppe.
Vanda wurde sofort von dem dortigen Alpha-Weibchen, Dua, angegriffen und
floh, im Fliehen noch die ihr bekannte Vau angreifend. Anschließend griffen
auch Mecki und Omega Vanda an, wobei sich Vanda nur gegenüber Omega
wehrte (einmal). Bereits am Abend des Introduktionstages hatte sie jedoch die
vierte Position in der Gruppe vor Omega erlangt. In den folgenden Tagen war
auffällig, dass sie besonders von Mecki angegriffen wurde und dass sie dem
Alpha-Weibchen, Dua, präsentierte und von diesem auch inspiziert wurde.
Bereits nach vier Tagen hatte sie ein besonderes Verhältnis zu den beiden
ranghohen Weibchen hergestellt und erreichte am 5. Tag nach Introduktion die
dritte Position in der Gruppe, die jedoch von Mecki nicht unangefochten
blieb.
Diese labile dritte Position konnte sie in den nächsten zwei Monaten nicht verbessern,
dies änderte sich auch nicht nach Introduktion ihrer Tochter Frieda (30.03.1976),
obwohl diese in der großen Nikita-Gruppe die erste Position innerhalb der Weibchen
trotz des Fehlens ihrer Mutter gehalten hatte, hier fiel sie auf den nach ihrer Mutter
nächsten Rang ab.
Kurzfristige Introduktion von Nikita in die Weibchengruppe Erst als wir kurzzeitig Nikita am 22.04. 1976 in die Weibchengruppe introduzierten, traten Änderungen auf. Nikita inspizierte und bestieg alle Weibchen, wobei Dua ihm auswich, dann aber das Männchen aufsteigen ließ und präsentierte. Der relativ friedliche Verlauf änderte sich schlagartig als Nikita kurz hintereinander Stirni und Mecki, letztere zu dem Zeitpunkt gerade geringfügig höherrangig als Vanda, angriff und biss. Vanda stürzte sich sofort auf beide, dabei Nikita präsentierend, und fügte Stirni mehrere ernste Bissverletzungen zu, die uns veranlassten, dieses Weibchen kurzfristig aus der Gruppe zu nehmen. Anschließend richtete sich Vandas agonistisches Aktivität gegen Mecki, wobei sie jedoch stets von Dua angegriffen wurde. Bei Zulassen von Stirni war diese wiederum Hauptadressat des agonistischen Verhaltens von Vanda. Auch nach erfolgter Absperrung von Nikita behielt Vanda die zweite Position in der Gruppe und entsprechend Frieda die dritte Position inne.
Reintroduktion von Vau und Introduktion von Omega Anschließend an den letzten Versuch wurden am 26.04.1976 Vau und am 27.04.1976 Omega in die große Gruppe introduziert. Beide Weibchen ließen sich ohne zu fliehen von Nikita inspizieren und besteigen und präsentierten bei seiner Annäherung sofort. Die reintroduzierte Vau näherte sich anschließend gleich den anderen ihr bereits bekannten Weibchen, Omega hingegen hielt anfänglich Abstand, ließ sich aber auch problemlos integrieren.
Fusion beider Gruppen Am 24.05.1976 fusionierten wir schließlich beide Gruppen. Es war auffällig, dass Vanda alle Weibchen der anderen (alten) Gruppe angriff, mit Ausnahme der erst relativ kurz aus ihrem Bereich entfernten Omega. Vornehmlich richtete sie ihre Aktivität gegen die im mittleren Bereich sich befindenden Weibchen, Blonda und Alba, weiterhin griff sie auch Dua an und sicherte sich die Alpha-Position der Gesamtgruppe.19 Während die drei ranghöchsten Weibchen der erweiterten Nikita-Gruppe sich agonistischer Aktionen enthielten, griff Alba mehrfach Stirni an und verwies sie somit auf einen niedrigeren Rang, entsprechend dürfte auch der Angriff Barbaras auf Dua ihren höheren Status bedingen.
Die beschriebenen besonderen Funktionen des Alpha-Männchens, Nikita, und des Alpha-Weibchens, Vanda, bei Introduktionen können sicher auf ihre Alpha-Positionen zurückgeführt werden. Auch im normalen Zusammenleben unserer Makaken-Gruppe, die zum Zeitpunkt dieser Experimente auf dreißig Tiere angewachsen war, zeichneten sich Alpha-Männchen und Alpha-Weibchen durch besonderes Verhalten aus:
Das Verhalten der Alpha-Tiere
Beide Alpha-Tiere übernehmen Verteidigungsaufgaben: So verlassen z. B. bei Flucht
unserer Makakenherde in die Außengehege beide als letzte rückwärtsgehend, dabei
auch oft biped aufgerichtet, die Innenkäfige, der Störung hierbei drohend, bis das
letzte Gruppenmitglied vor dieser geflüchtet ist. Gleichfalls betreten sie auch wieder als
erste die Innenkäfige, wobei sich Nikita weiter vorwagt. Intensiviert wird ihre
Verteidigungsbereitschaft, wenn Jungtiere in den Innenkäfigen zurückgeblieben sind,
dann springen sie drohend an die Vorderfront des Käfigs und greifen durch das Gitter,
so den Jungtieren den Rückzug sichernd.
Beide Alpha-Tiere haben bei der Nahrungsaufnahme absolute Priorität, wobei Nikita
wiederum die Priorität vor Vanda hat, wenngleich Vanda unbedroht gleichzeitig mit Nikita
essen darf. Alle anderen Tiere hingegen warten die Nahrungsaufnahme durch Nikita
ab.20
Beide Alpha-Tiere zeigen gegenüber anderen Gruppenmitgliedern das Verhalten des Drohens mit offenen Mund, bei Nikita oft verbunden mit Grunzlauten (stummes Drohen, bzw. Drohen mit Grunzen ([2])) . Beide Alpha-Tiere können jederzeit jedes andere Tier der Gruppe putzen, werden jedoch selber in der Regel nur nach Aufforderung geputzt.
Das Verhalten des Alpha-Männchens
Nikita bedroht bei Gefahr alle Gruppenmitglieder, die sich von der Gruppe entfernen,
was an Hüteverhalten bei Papio hamadryas erinnert, er besteigt bei Bedrohung
sukzessive alle Weibchen der Gruppe, insbesondere diejenjgen, die Fluchtintentionen
zeigen, was wiederum an Besitzsicherung bei Mantelpavianen Papio hamadryas
erinnert.
Nikita interveniert bei agonistischen Interaktionen, meist bedroht er den
rangniedrigeren der beiden Kontrahenten, desgleichen bedroht er auch häufig
rangniedrige Tiere, die sich ranghohen nähern.
Nikita beißt und verfolgt Weibchen, die vor ihm fliehen, bzw. nach Aufsteigen vor ihm
Fluchtintentionen zeigen, er bedroht ältere männliche Jungtiere, die bei adulten
Weibchen aufsteigen und steigt häufig nach Verjagen der Jungtiere selber auf.
Weiterhin bedroht er bei nicht vorhandener Spielappetenz auch Jungtiere, die sich ihm
zu weit nähern, bzw. ihn zufällig berühren.
Nikita zeigt positive soziale Aktivität (soziale Körperpflege, Spiel) gegenüber den
Weibchen meist nach Aufsteigen und mehrfach am Tage gegenüber den Jungtieren.
Seine Spielappetenz äußerst sich durch kurze Hustlaute, verbunden mit tiefem
Brummen, Vorschieben der Mundpartie und - hin und wieder - Hochziehen der Stirn.
Bei den Spielen dürfen die Jungtiere auf ihm herumkrabbeln, ihn beißen;
er wiederum zieht diese an sich, zerrt an ihnen und beißt spielerisch nach
ihnen.21
Das Verhalten des Alpha-Weibchens
Vanda sitzt in der Regel ruhig, scheinbar teilnahmslos auf einem erhöhten Ruheplatz,
beobachtet dabei jedoch stets die übrige Gruppe.
Vanda flieht im Gegensatz zu den anderen Tieren nur sehr selten vor Nikita, in
der Regel geht sie nur langsam beiseite oder wendet das Gesicht ab, dabei
häufig präsentierend. Desgleichen präsentiert sie ständig bei Drohen gegenüber
anderen Gruppenmitgliedern (gesichertes Drohen); wohl mit dieser permanenten
Besänftigung verbunden, beschränken sich die sexuellen Interaktionen zwischen
Nikita und Vanda auf Inspizieren und Aufsteigen ohne Intromission. So ist
auch Vanda das einzige Weibchen der Gruppe, das nicht jährlich ein Junges
bekommt.
Vanda verdrängt häufig andere Tiere von deren Platz, indem sie nach ihnen greift
oder an ihnen zerrt. Bei Pressen der Weibchen konnten wir wiederholt beobachten,
dass Vanda von hinten auf die Weibchen kletterte und sich sogar auf deren Kopf
setzte.
Vanda beißt Tiere, die vor anderen fliehen, dabei bleibt sie in der Regel sitzen, bzw.
beugt sich nur mit dem Kopf und dem Oberkörper nach vorn.
Vanda vermeidet bei dem friedlichen Näherrücken den direkten Sichtkontakt, indem
sie nach oben blickt, bzw. nähert sich auch anderen Weibchen präsentierend.
Entsprechendes beobachteten wir auch bei dem Zusammenlassen von Stirni mit der
rangniedrigeren Omega.
Vanda sucht vornehmlich Weibchen, die Jungtiere mit sich herumtragen, auf, putzt
diese, umgreift sie von hinten, bzw. versucht besonders rangniedrigen Weibchen die
Jungen zu entreißen. „Gestohlene“ Jungtiere trägt sie mit sich herum und putzt
sie. Das gleiche gilt für frei sich bewegende Jungtiere, nach denen sie häufig
greift.
Bei Macaca fascicularis ist - wie auch bei anderen Makakenspecies - die Rangfolge der
Weibchen linear (vgl. aber Unterkapitel 14.9). Durch unsere Untersuchungen haben wir
aufgezeigt, dass diese lineare Reihenfolge auf komplexen Beziehungen beruht. Es gibt
wohl zwei verschiedene Gruppen von Weibchen, nämlich potentiell ranghohe
und potentiell rangniedrige (z. B. Vau und Omega). „Ranghohe“ Weibchen
haben das Bestreben, ihre soziale Position zu verbessern bzw. zu sichern,
„rangniedrige“ hingegen sind unsicher und erreichen trotz bester Vorbedingungen (so
wurde Vema als dritte introduziert) keine höhere Position in der Gruppe, da sie
keine Versuche unternehmen, ihren Rang zu verbessern, und bei agonistischen
Interaktionen mit anderen Gruppenmitgliedern schnell aufgeben. Neben diesem
„natürlichen“ Rang scheint der Rang des Individuums abhängig zu sein von dem
Zeitpunkt der Introduktion (früher introduzierte Tiere haben einen Platzvorteil vor
später introduzierten), von den Beziehungen zu anderen Gruppenmitgliedern
und schließlich auch von dem Verhalten des Individuums bei Erstbegegnung,
nämlich dem möglichst frühen Schaffen einer guten Ausgangsposition für weitere
Gruppenauseinandersetzungen. Hier ist auch ein Unterschied im Verhalten
ranghöherer und rangniedriger Weibchen zu erkennen: während ranghöhere ihre
Stellung sofort sichern, sind rangniedrigere unsicher.
Darüber hinaus sind verwandtschaftliche Beziehungen in Freiland-Gruppen
und in zunehmendem Maße auch in unserer Kolonie für den späteren Rang
entscheidend. Das „Vererben“ des Ranges auf die Jungtiere, wurde in diesem
Unterkapitel für Frieda aufgezeigt, die auch bei Abwesenheit der Mutter schon im
Alter von einem Jahr die höchste Rangposition in der Nikita-Gruppe hielt.
Entsprechendes konnten wir später bei weiteren Jungtieren beobachten (s.
u.).
Bemerkt werden soll schließlich noch, dass der Oestrus eines Weibchens zu keiner
Rangverbesserung führt.
Allen Weibchen war gemeinsam, dass sie die gegen sie gerichteten agonistischen
Aktivitäten durch Einnehmen einer Demutshaltung, durch Verzicht auf Flucht und
Gegenwehr verringerten und durch Präsentieren sowohl dem Männchen als auch den
Weibchen gegenüber die Tendenz zu einem Stimmungsumschwung verstärkten. Diese
Strategie der Vermeidung agonistischer Auseinandersetzungen wurde unabhängig vom
späteren Rang angewandt, bei Jungfrau nur mit großer zeitlicher Verzögerung. Das
nächste Ziel war dann, zu nächstrangigen Individuen Beziehungen gegenseitiger
sozialer Körperpflege herzustellen. Gelang diese „Freundschaftsstrategie“ wurde die
einmal erreichte Position durch gemeinsames Bedrohen rangniedrigerer Tiere
gesichert, bzw. durch Wiederholen der Strategie konsequent ausgebaut, indem
nun wiederum das vorher gleichrangige Tier bedroht wurde. Hierbei können
auch mehrere Rangstufen gleichzeitig übersprungen werden. So bildete z.B.
Barbara zuerst zu Jungfrau besondere Beziehungen aus, dann zu Zita, so Alba
überspringend.
Bei dem zum Rangaufstieg notwendigen Einsatz des Individuums muss dieses seine
eigene soziale Position richtig einschätzen, um Mehrfrontenkämpfe, verbunden mit
hoher Verletzungsgefahr, zu vermeiden. Die einmal erreichte Position wird anfangs
auch durch erhöhte agonistische Aktivität gegenüber den rangniedrigeren
verstärkt. Dies ist besonders auffallend bei rangmittleren Weibchen, ihre höhere
agonstische Aktivität ist wohl bedingt durch ihre labile Position in der Gruppe.
Weiterhin ist für den Rangaufstieg wichtig, dass die Weibchen gegebene Chancen
konsequent nutzen. Hier dürfte der individuellen Fitness besondere Bedeutung
zukommen.
Das Beispiel perfekt angewandter Strategien zeigte Vanda. Bei ihrer Introduktion in
die Weibchengruppe vermied sie jegliche agonistische Auseinandersetzungen mit ihr
fremden Tieren, bzw. ertrug gegen sie gerichtete Aktivitäten passiv, dabei jedoch
den höheren Rang gegenüber der ihr bekannten Vau sichernd. Nach erfolgter
Beruhigung in der Gruppe erreichte sie durch gezieltes Drohen gegenüber
der rangniedrigen Omega eine höhere Position. Danach stellte sie besondere
Beziehungen gegenseitiger sozialer Körperpflege zu den beiden ranghöchsten
Individuen (Dua, Stirni) her, so tendenziell Mecki überspringend. Ihre labile dritte
Position versuchte sie nicht auszubauen. Erst als der kurzfristig introduzierte
Nikita Stirni angriff, reagierte sie sofort, griff das Weibchen an und verstärkte
ihre einmal erreichte Rangposition durch unzählige Bisse. Den nun erlangten
zweiten Rang konnte sie nicht verbessern. Schließlich richtete sie bei der Fusion
der beiden Gruppen ihre agonistischen Aktivitäten vornehmlich gegen die
wegen deren labiler Position besonders „gefährlichen“ rangmittleren Weibchen,
während sie die ranghöheren, inclusive Dua, auf eine rangniedrigere Position
verwies.
Zusammenfassend konnten wir durch Introduktionen, Reintroduktionen und
Fusionsexperimente mit 16 Macaca fascicularis (1 Männchen, 15 Weibchen)
zeigen:
1. Bei Introduktionen kommen dem Alpha-Männchen und dem Alpha-Weibchen
besondere Funktionen zu, was ihrem Verhalten in der Gruppe entspricht.
2. Die Beziehungen zwischen Alpha-Männchen und neuintroduziertem Tier verlaufen
nach einer festen Regel mit verschiedenen Stadien der Integration. Auf die 1. Stufe
„Verfolgen“ (durch das Männchen), folgt die 2. Stufe „Präsentieren“ (des Weibchens),
die 3. Stufe „Besteigen“ (durch das Männchen), die 4. Stufe „Beißen in den
Rücken“ (durch das Männchen) und schließlich die 5. Stufe „gegenseitige soziale
Körperpflege“. Die gleiche Regel gilt auch für die dyadischen Beziehungen in der
Nur-Weibchen-Gruppe.
3. Das Alpha-Weibchen beißt gleich zu Beginn der Introduktion - vor oder nach dem
Männchen - das neuintroduzierte Tier.
4. Bei den Weibchen scheint es im Sinne eines natürlichen Ranges ranghohe und
rangniedrige Individuen zu geben. Der spätere Rang ist darüber hinaus abhängig von
unterschiedlichen Faktoren, wie Zeitpunkt der Introduktion und Beziehungen zu
anderen Individuen.
5. Weibchen wenden zur Erreichung eines höheren Ranges bestimmte Strategien an,
wobei zwei grundsätzliche Strategien erkannt wurden, die Strategie der Vermeidung
agonistischer Auseinandersetzungen und die Strategie der Herstellung von Beziehungen
gegenseitiger sozialer Körperpflege zu ranghöheren Tieren.
Bei der experimentellen Gruppenbildung22 unserer Javanermakakenkolonie in den Jahren 1975 bis 1976 (vgl. 14.2) schien es so, als gäbe es im Sinne eines natürlichen Ranges hochrangige und niederrangige Individuen. Diese unterschieden sich in ihrem Verhalten während der Introduktionsversuche voneinander. Der endgültige Rang hing zusätzlich von verschiedenen Faktoren ab, wie vom Zeitpunkt der Introduktion und von Beziehungen zu anderen Tieren der Gruppe. Ziel vorliegender Untersuchung war es daher, durch geeignete Versuchsanordnungen zu klären, inwieweit der endgültige Rang mehr oder weniger zufällig ist, oder aber ob der Rang als eine Eigenschaft des Individuums angesehen werden kann. Ist ein Rangplatz von Zufällen abhängig, dann dürfte die frühere soziale Erfahrung die Rangordnung in einer neu etablierten Gruppe nur wenig beeinflussen. Wäre dagegen der Rang eine Eigenschaft des Individuums, dann müsste unser Wissen über den relativen Rang eines Individuums Voraussagen über den späteren Rang unter neuen sozialen Bedingungen zulassen.23 Neben der auffälligen linearen Dominanzhierarchie sind Verwandtschaftsbeziehungen („kinship-relations“) von hoher Bedeutung für die Makakensozialstruktur (vgl. 14.4). Zudem wollten wir nach Gesetzmäßigkeiten suchen, die die sozialen Beziehungen in der Gruppe regeln.24
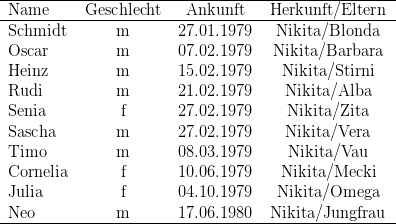
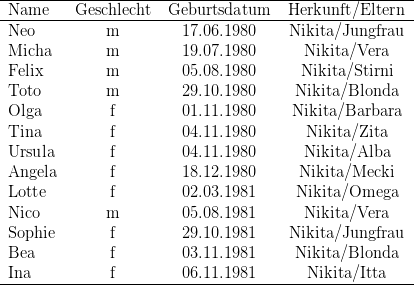
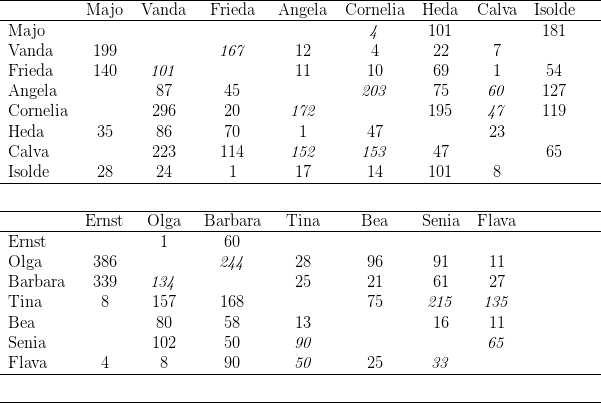
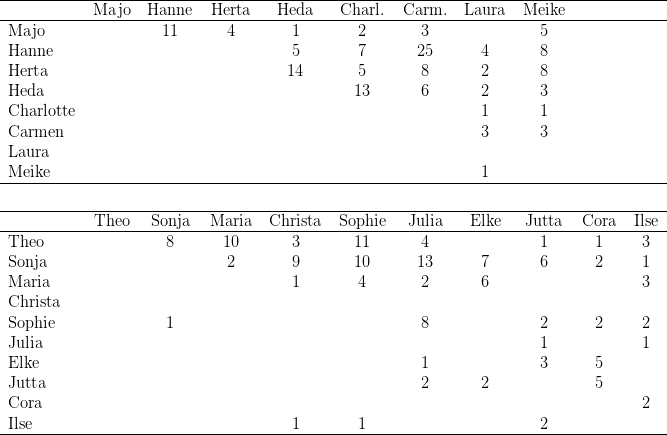
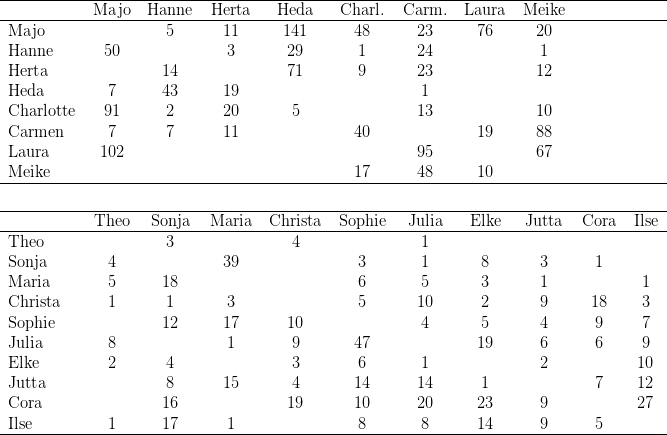
Zwölf der zu Versuchsbeginn in Kassel gehaltenen 29 Macaca fascicularis waren Wildfänge, die restlichen 17 wurden in den letzten Jahren in Kassel geboren. Während der Untersuchung nahm die Gruppe durch weitere Geburten um neun Tiere zu; ein Weibchen starb an Geburtskomplikationen; so dass die Gruppe zu Ende der Untersuchungen aus 37 Individuen bestand. Bei der Auswertung wurden die Daten derjenigen Tiere berücksichtigt, die während der gesamten Untersuchung im Gruppenverband anwesend waren. Zudem verzichteten wir auf die Darstellung der Ergebnisse dreier „mutterloser“ Individuen (Birgit, Benni, Vemo). Die vorgestellten Daten beziehen sich also auf 25 der 28 Individuen (vgl. Tabelle 14.2)
Die Haltungsbedingungen sind in Unterkapitel 14.2.1 beschrieben. Zusätzlich waren für indirekte Beobachtungen drei Videokameras fest montiert, zwei Kameras lieferten Übersichtsbilder über den Haltungskäfig 1 bzw. die Haltungskäfige 2 a bis 2 c; die dritte war beweglich und steuerbar (Schwenk- und Neigekopf, Zoom, Blende, Scharfeinstellung).
Die Interaktionen der Tiere wurden täglich drei Stunden durch zwei von sechs Beobachtern protokolliert. Dabei achteten wir darauf, dass jede Stunde der Aktivitätszeit (7.00 bis 19.00 Uhr) dreimal berücksichtigt war und verschoben hierzu den Beginn der im Block stattfindenden dreistündigen Beobachtungen täglich um eine Stunde. Im einzelnen protokollierten wir die Interaktionen der Gesamtgruppe (ein Beobachter 2,5 Stunden) und einzelner Individuen (ein Beobachter 0,5 Stunden).25
Nach jedem Experimentalschritt, der gewöhnlich zwölf Tage dauerte, führten wir einen Futterkonkurrenztest durch. Hierbei wurde jedes der adulten Weibchen sukzessive mit jedem anderen adulten Weibchen der Gruppe abgesperrt. Das Vorgehen ist in Unterkapitel 14.2.1 bereits beschrieben.
Die bei Abschluss der Gruppenbildung (Mai 1976) entstandene
Rangordnung blieb im Wesentlichen stabil und änderte sich auch
nicht im Zusammenhang mit Geburten und dem Heranwachsen von
Jungtieren.26
Um zu prüfen, inwieweit die Ränge und damit die sozialen Positionen der einzelnen
Individuen abhängig von besonderen Beziehungen zu anderen Individuen sind, führten
wir von Januar bis Mai 1979 einige Separations- und Reintroduktionsexperimente
durch (E.S. 1.0 bis 2.1).
Wir separierten das Alpha-Männchen (E.S. 1.1), das Alpha-Weibchen (E.S.
1.3) und das Beta-Weibchen (E.S. 1.5), bzw. alle drei Individuen gemeinsam
(E.S. 1.7) und reintroduzierten sie in die Gruppe (E.S. 1.2, 1.4, 1.6, 1.8, 1.9,
1.10). Zudem separierten wir drei ranghöhere (E.S. 1.11) und drei rangniedrige
Clane (E.S. 2.1). Offensichtliches Ergebnis dieser Versuchsserie war nun, dass
die Abwesenheit der jeweils separierten Individuen für die Rangstruktur der
Gruppe keine Bedeutung hatte. Der Gruppenzusammenhalt blieb auch ohne
Alpha-Männchen, Alpha-Weibchen und Beta-Weibchen erhalten, die Häufigkeit
agonistischer Interaktionen nahm nicht zu, vielmehr erhöhten sich nach der
Separierung der beiden ranghöchsten Weibchen die positiv-sozialen Interaktionen
sogar, was aber nicht zwingend das Resultat der Separation war, sondern
eventuell auch auf die abgeschlossene Geburtensaison zurückgeführt werden
kann.
Auffällige Änderungen beobachteten wir bei juvenilen Männchen, zeigten diese doch erst
nach der Separation von Nikita, dem Alpha-Männchen, untereinander agonistisches
Verhalten.27
Durch ihr Verhalten bei der Abwesenheit von Nikita sicherten sich die
Männchen untereinander offensichtlich den Rang der jeweiligen Mutter,
dabei gingen sie äußerst „sparsam“ vor (s. o.) und bedrohten in der Regel
nur Söhne von solchen Müttern, die ihren eigenen Müttern gegenüber
nächstrangig waren. Desweiteren versuchten die juvenilen Männchen nun die
Weibchen zu kontrollieren, hierdurch die „Aufgaben“ des Alpha-Männchens
übernehmend.28
Die juvenilen Männchen beschränkten sich dabei auffällig auf Weibchen, die
rangniedriger waren als ihre eigenen Mütter.
Wir können hier nicht entscheiden, ob das Verhalten der Weibchen, gerade mit
rangbenachbarten Individuen enge soziale Kontakte zu unterhalten, für das
Aufrechterhalten der sozialen Bindungen verantwortlich war oder ob das Verhalten der
juvenilen Männchen als der Faktor der Gruppenstabilität gelten kann, dass
also ohne diese die Gruppenstruktur verändert worden wäre. Doch bleibt
auch bei Annahme einer solchen Funktion juveniler Männchen das Ergebnis
unerwartet: Die sozialen Beziehungen sind konstant, auch wenn die Tiere,
die eindeutig und unbestreitbar das Zusammenleben steuern und regulieren,
abwesend sind. Sichtbar hielt das Netzwerk sozialer Beziehungen, das die Tiere in
den vorausgegangenen Jahren aufbauen konnten, auch solchen Belastungen
stand.
Daher versuchten wir durch geeignete Versuchsanordnungen, dieses feste
Gruppengefüge zu lösen. Da die sozialen Beziehungen der einzelnen Gruppenmitglieder
vor allem auf Rangbeziehungen beruhten, sollten die Ränge der einzelnen Individuen
experimentell geändert werden. Durch diese Versuche konnte gleichzeitig die
Frage nach einer eventuellen Disposition experimentell angegangen werden.

Von unseren Introduktionsexperimenten bei Gruppenbildung wussten wir, dass der Introduktionszeitpunkt von hoher Bedeutung für die spätere soziale Stellung des Individuums ist, dass es für später hinzukommende Tiere weitaus schwieriger ist als für die bisherigen Gehegebewohner, sich in der Gruppe durchzusetzen. Daher beschlossen wir, den rangniedrigen Clanen Platzvorteile zu geben, so ihnen experimentell bessere Startchancen verschaffend. Tiere, die hierdurch ranghoch wurden, bezeichnen wir im folgenden als „künstlich ranghohe“ Individuen.
Bildung der Ausgangsgruppe und erste Introduktionsversuche
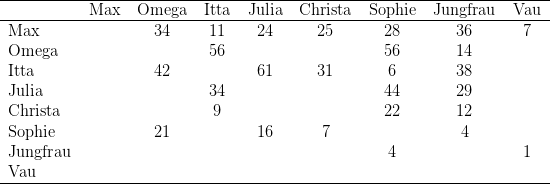
Zu Beginn dieser Experimente separierten wir die drei rangniedrigsten Clane (E.S. 2.1)
und hielten diese 14 Tage gemeinsam in den Käfigen 2-4 des Haltungsraumes. In dieser
Gruppe war Stirni eindeutig ranghöchstes Weibchen, ihre Tochter Itta hielt
unangefochten die 2. Rangposition. Agonistische Interaktionen zwischen Vau und
Omega konnten nicht beobachtet werden. Doch ergab der Futterkonkurrenztest zu
Ende der 14 Tage, dass Vau eindeutig ranghöher als Omega war, dass also die
Rangbeziehungen in der Ausgangsgruppe unverändert erhalten blieben.
Bei den nun im vierzehntägigen Abstand folgenden Introduktionen der übrigen Clane
entgegengesetzt zur Rangfolge war auffälligstes Ergebnis, dass die ehemals ranghöheren
Weibchen von den drei vorher rangniedrigsten Clanen der Ausgangsgruppe
angegriffen, gebissen und gejagt wurden; dies beobachteten wir bei der Introduktion
von Jungfrau (E.S. 2.2), Alba (E.S. 2.3), Barbara (E.S. 2.4), Zita (E.S. 2.5,
Blonda (E.S. 2.6) und Vera (E.S. 2.7). Stirni und Itta erreichten hierdurch
stets unbestritten die höchsten Rangpositionen in der Gruppe, waren also
(wenn auch „künstlich“) Alpha- und Beta-Weibchen und vom Verhalten her
vergleichbar mit Vanda und Frieda in der Gesamtgruppe. Entsprechend konnten
auch Vau und Omega ihren höheren Rang halten, wenngleich Omega den
dritten Rangplatz vorübergehend an Jungfrau verlor. Doch waren die „künstlich
ranghohen“ Weibchen nicht in der Lage, sich auch gegenüber dem juvenilen Männchen
Berni durchzusetzen. Vau und Omega gelang dies zudem auch nicht bei Tritus und
Ernst.
Theoretisch hätte es nun sein können, dass die resultierende neue Rangordnung sich
genau entgegengesetzt zur alten (in der Gesamtgruppe) etablieren würde, dass
also die jeweils zuletzt introduzierten Tiere auch die niedrigsten Positionen
einnehmen würden, doch war dies auffällig nicht der Fall. Einige Weibchen,
Barbara, Zita, Blonda und Vera, konnten sich zwar gegenüber den drei „künstlich
ranghohen“ Clanen und gegenüber der erstintroduzierten Jungfrau nicht durchsetzen,
bedrohten aber die vor ihnen Introduzierten (Barbara: Alba (E.S. 2.4), Zita:
Barbara und Alba (E.S. 2.5), Blonda: Barbara und Alba, nicht jedoch Zita
(s. u.) und Vera schließlich Blonda, Barbara und Alba). Sie sicherten sich
gegenüber den von ihnen bedrohten Tieren ihre ursprünglich höheren Positionen
in der Gruppe, ein Verhalten, das wir „Rangsicherungsverhalten“ genannt
haben.29
Eine ganz andere Qualität hatte die Introduktion des Mecki-Clanes, Mecki
wurde zwar gleich nach Betreten des Käfige ebenfalls von den drei „künstlich
ranghohen“ Weibchen und deren Kindern angegriffen, konnte sich jedoch in einem ca.
einstündigen Kampf behaupten und die Position des Alpha-Weibchens in der
neuen Gruppe einnehmen. Dabei bedrohte sie alle anderen Weibchen der
Gruppe und deren Kinder (E.S. 2.8). Nämliches galt für ihren Sohn Majo und
eingeschränkt für ihre Tochter Calva, die sich gegenüber Berni und Orbi, den zwei
ältesten juvenilen Männchen nicht durchsetzte. Im Zuge dieser agonistischen
Auseinandersetzungen gelang es den adulten Weibchen, die vorher auch das
„Rangsicherungsverhalten“ gezeigt hatten, nun ihren Rang zu verbessern, sie
bedrohten die „künstlich Ranghohen“ und sicherten sich hierdurch mehr oder weniger
ihre alten Positionen. Barbara bedrohte zudem auch Zita und tauschte mit
ihrem ehemaligen Rangpartner die Position, Nämliches galt für Blonda und
Vera.
Auffälligerweise zeigten jedoch Jungfrau und Alba das „Rangverbesserungsverhalten“ nicht,
entsprechend behielten diese auch im Vergleich zu Stirni, Vau und Omega ihre
niedrigeren Rangpositionen. Alba stellte nur im Verhältnis zur vorher Rangnächsten,
Jungfrau, die alte Rangfolge wieder her. Die Introduktionen von Frieda, Vanda und
Nikita änderten an dem Ranggefüge nichts mehr. Nach ihren Introduktionen
nahmen alle drei jeweils unbedroht und unangefochten ihre alten Positionen
ein.
Offensichtlich gibt es also - wie in Unterkapitel 14.1 bereits betont - Weibchen
unterschiedlicher Qualität, 1. Weibchen, die auch in neuen Situationen ihre alten
Positionen behaupten können, wollen wir als “ranghoch“ bezeichnen, 2. Weibchen, die
sich zwar in neuen Situationen nicht durchsetzen können, jedoch versuchen, ihre alten
Rangbeziehungen gerade zum ehemals Rangnächsten zu sichern und darüber hinaus
offensichtlich auf die Chance warten, ihren Rang in sozialen Konfliktsituationen zu
verbessern, wollen wir als „rangmittel“ bezeichnen, 3. Weibchen, die weder
„Rangsicherungs-“ noch „Rangverbesserungsverhalten“ zeigen, als „rangniedrig“.

Kontrollversuch
Eine mögliche Erklärung des bisherigen Versuchsergebnisses hätte sein können,
dass es sich bei Stirni in Wirklichkeit um gar kein rangniedriges Weibchen
handelte, dass sie in Wirklichkeit ranghoch war und nur durch negativ-soziale
Beziehungen zu den ranghöchsten Individuen der Gruppe einen niedrigeren
Rangplatz in der Hierarchie einnahm. Um dies auszuschließen, führten wir einen
Kontrollversuch durch (E.S. 3.0 bis 3.8), bei dem wir erneut drei rangniedrige Clane
separierten, diesmal jedoch ohne Stirni und Itta. Stattdessen sperrten wir zusätzlich
zu Vau/Christa und Omega/Paul nun Jungfrau und ihren Sohn Protus ab.
Jungfrau war zu diesem Zeitpunkt eindeutig rangniedrigstes Weibchen in der
Gesamtgruppe.
In der neuen Ausgangsgruppe blieben wiederum die alten Rangbeziehungen erhalten.
Vau war ranghöchstes Weibchen, ihre Tochter Christa hielt die zweite Position.
Nach vierzehntägiger gemeinsamer Haltung introduzierten wir dann erneut
sukzessive die übrigen Clane entgegengesetzt zur nun in der Gesamtgruppe
bestehenden Rangordnung (Stirni introduzierten wir jedoch aus oben genannten
Gründen. abweichend vom generellen Versuchsaufbau, als letztes Weibchen vor
Mecki).
Augenscheinlichstes Ergebnis des gesamten Kontrollversuchs war nun, dass wieder die
drei adulten Weibchen der Ausgangsgruppe die höchsten Rangpositionen hielten. Auch
zeigten diesmal Zita, Barbara, Vera und Blonda „Rangsicherungsverhalten“, nicht
jedoch Alba und Stirni. Zita gelang es zudem, genauso wie bei den vorangegangenen
Experimenten, besondere Beziehungen zu den ranghöchsten Weibchen (den „künstlich
Ranghohen“) aufzubauen; sie erlangte hierdurch in der neuen Gruppe eine
relativ hohe Rangposition. Da die Introduktion des Mecki-Clanes die nämlichen
Ergebnisse erbrachte wie bei dem vorangegangenen Versuch, - Mecki war
unbestritten ranghöchstes Weibchen, die rangmittleren Weibchen begannen
die „künstlich Ranghohen“ zu bedrohen - brachen wir diesen Versuch nach
dreißig Minuten ab, sperrten den Mecki-Clan wieder zu Nikita/Vanda/Frieda
und hielten die übrigen Tiere in unveränderter Zusammensetzung. In den
nächsten vier Wochen änderte sich an den sozialen Beziehungen nichts mehr. Die
bisher introduzierten adulten Weibchen und ihre Kinder waren offensichtlich
nicht in der Lage, ohne „Anstoß“ von außen ihre ursprüngliche Position zu
erreichen.
Weitere Introduktionsversuche
Aus der Gruppe, die aus Weibchen rangniedriger und rangmittlerer Clane bestand und
in der die rangniedrigen Clane „künstlich ranghoch“ waren, separierten wir daher die
„künstlich Ranghohen“. Durch den Wegfall dieser Individuen änderte sich nur wenig,
die übrigen Tiere behielten mehr oder weniger ihre bisherige Position, Rangwechsel
fanden nur zwischen Blonda/Zita und Stirni/Alba statt. Damit stellten Stirni und
Blonda jeweils die ursprünglichen Situationen in der Gesamtgruppe wieder
her.
Anschließend introduzierten wir dann sukzessive die „künstlich Ranghohen“ entgegengesetzt
zur Rangfolge. Nun war keines der „künstlich ranghohen“ Individuen in der Lage, sich in der
Gruppe, in der sie vormals unangefochten die höchsten Ränge gehalten hatte, zu behaupten.
Die „künstlich Ranghohen“ mussten sich vielmehr mit den untersten Rangplätzen zufrieden
geben.30
Die Introduktion von Mecki war diesmal qualitativ verschieden von den vorherigen,
zwar schaffte es Mecki während der ersten drei Stunden nach Introduktion mühelos,
die anderen adulten Weibchen zu kontrollieren, doch konnte sie sich nicht gegenüber
Berni und Orbi durchsetzen. Dies könnte auch der Grund sein, warum Mecki ca. sechs
Stunden nach Introduktion wohl auch von adulten Weibchen gebissen und
niedergekämpft wurde, versuchsplanbedingt haben wir diese Interaktionen nicht
beobachtet. Nach Aussagen unserer Tierpfleger hat sich ein ganzer Pulk von
Gruppenmitgliedern wohl gleichzeitig auf Mecki gestürzt. Wir sahen nur das Ergebnis,
Mecki saß schwer verletzt in einer Käfigecke am Boden, ihre jüngste Tochter an sich
gepreßt und in engem Körperkontakt mit ihrer ebenfalls verletzten Tochter
Calva.31
Hierdurch sank Mecki im Rang; bei Futterkonkurrenztests zeigte sie nur gegenüber
Jungfrau agonistische Reaktionen. Nun war Blonda unbestritten ranghöchstes
Weibchen und Berni, ihr ältester Sohn, ranghöchstes Männchen, Orbi zweitrangiges
Männchen.
Da die anschließenden Introduktionen von Frieda, Vanda und Nikita ohne
agonistische Auseinandersetzungen erfolgten, diese drei also erneut unbestritten die
ranghöchsten Positionen einnahmen, gab es auch keine Chance für Mecki, ihren Rang
schlagartig zu verbessern. Sie verbesserte diesen vielmehr Schritt für Schritt.

Es sei hier bemerkt, dass Berni, Orbi und Majo bei Friedas Introduktion versuchten,
diese zu inspizieren und zu besteigen und somit zu kontrollieren. Frieda nahm jedoch
offensichtlich kaum Notiz von den jungen Männchen und bedrohte unabhängig von
dem, was „hinter ihr geschah“, adulte Weibchen. Wurde sie von einem der
Männchen bestiegen, drehte sie sich leicht um und führte mit ihrer Hand einen
Schlag gegen den Kopf oder Körper des Aufsteigers. Teilweise führte dies
dazu, dass die juvenilen Männchen sich nicht mehr auf dem Brett halten
konnten.
Offensichtlich waren alle unsere Individuen in der Lage, einen hohen Rang zu halten.
Der Rang eines Individuums in einer bestehenden Gruppe ist somit relativ und vor
allem von den anderen Gruppenmitgliedern abhängig. Doch gibt es nach unseren
Befunden Gruppenmitglieder unterschiedlicher Qualität, deren Qualität wir mit
ranghoch, rangmittel und rangniedrig umschrieben haben. Der Rang wäre demnach ein
Maßstab der sozialen Fitness und Eigenschaft des Individuums, was natürlich nicht
zwangsläufig bedeutet, dass jedes Individuum in jeder sozialen Situation auch seinen
Platz halten muss.
Beide Fragen, die nach dem Vorteil und die nach dem Reproduktionserfolg,
können insofern abschließend nicht beantwortet werden. Insgesamt gesehen
wäre der Vorteil wohl auf beiden Seiten, Ranghohe verteidigen die Gruppe,
Rangniedrige weisen (eher unbewußt) neue Wege. Durch deren zwangsläufig höhere
Mobilität könnten zumindest theoretisch neue Nahrungsressourcen und neue
Lebensräume für die Gruppe erschlossen werden. Wir haben zudem gezeigt, dass
zumindest in Menschenobhut bei der Bildung und bei dem Aufrechterhalten
der Rangordnung verwandtschaftliche Beziehungen von hoher Bedeutung
sind.
In unserer Kolonie32 können wir nach ihrem Verhalten sechs verschiedene Typen von Individuen unterscheiden, nämlich drei, die jeweils nur durch ein Individuum repräsentiert sind ((1) Alpha-Männchen, (2) Alpha-Weibchen und (3) Beta-Weibchen), (4) die adulten Weibchen mit Ausnahme des Alpha- und Beta-Weibchens, (5) die juvenilen Männchen und (6) die juvenilen Weibchen. Der grundsätzliche Befund, dass wir nach dem Verhalten verschiedene Typen von Individuen unterscheiden können, gilt wohl grundsätzlich für alle Makakenspecies. Dies dürfte vor allem daran liegen, dass Weibchen und Männchen generell unterschiedliches Verhalten zeigen.
Ziehen wir zusätzlich in Betracht, dass adulte Individuen andere Verhaltensmuster zeigen als juvenile und infantile, so wären 4 Typen von vornherein zu erwarten, nämlich 1. adulte Männchen, 2. adulte Weibchen, 3. juvenile Männchen und 4. juvenile Weibchen. Von diesen vier Typen waren zum Zeitpunkt der Datenerhebung für die hier vorgestellten Befunde nur drei in unserer Gruppe hinreichend vertreten, fehlten doch bedingt durch die Gruppenbildung aus einem adulten Männchen und mehreren adulten Weibchen (noch) weitere adulte Männchen. Von den drei weiteren von uns herausgestellten Typen wird nur der des Alpha-Männchens in der Literatur hinreichend berücksichtigt.
Das Verhalten des Alpha-Männchens haben wir bereits ausführlich beschrieben (vgl. Unterkapitel 14.2.2), ihm kommt ohne Zweifel die Führungsposition in der Gruppe zu, es allein kann alle anderen Individuen der Gruppe bedrohen, niemals war irgendein agonistisches Verhalten gegen es gerichtet. Während unserer Beobachtungen war das Alpha-Weibchen das einzige Tier der Gruppe, das regelmässig den Kontakt zum Alpha-Männchen suchte. Die übrigen Mitglieder der Gruppe verhielten sich ihm gegenüber passiv, Kontakte gingen fast ausschließlich vom Alpha-Männchen aus und waren - sehen wir einmal von solchen zu oestrischen Weibchen ab - vornehmlich agonistisch. Bei abnehmender Gruppengröße nahmen seine Interaktionen zu den jeweils noch in der Gruppe verbliebenen Weibchen zu. Geburten in der Gruppe beeinflussten sein generelles Verhalten nicht, doch war auffällig, dass es die an sich schon geringen Kontakte zu anderen Weibchen nach den Geburten noch weiter reduzierte. Sein agonistisches Verhalten richtete es vornehmlich gegen alle männlichen Jungtiere (Majo, Berni, Ernst, Orbi, Max, Protus, Paul) bzw. gegen die Mütter der ranghöchsten juvenilen Männchen (Majo, Berni), nämlich gegen die adulten Weibchen Mecki und Blonda. Die besondere Stellung des Alpha-Männchens wird für alle Makakengruppen im Freiland und im Labor beschrieben, ihm kommt eine Kontrollfunktion zu, durch seine Interventionen werden agonistische Auseinandersetzungen beendet; es verhindert hierdurch stärkere Verletzungen. Darüber hinaus ist das Alpha Männchen der Beschützer der Gruppe. Wie wir gezeigt haben, wird die Bedeutung des Alpha-Männchens der Javanermakaken für den Gruppenzusammenhalt jedoch häufig überschätzt. Besondere Bedeutung kommt ihm sicher bei der Gruppenneubildung, gerade im Labor, zu. In einer bestehenden Gruppe hingegen ändert sich bei seiner Abwesenheit nur wenig. Auch unterstützt das Alpha- Männchen der Javanermakaken nicht den Angegriffenen, sondern beteiligt sich auch zugunsten des Angreifers und bedroht gemeinsam mit diesem den Unterlegenen.
Durch seine Interventionen werden somit, wie de Waal 1978 betont, „die Dinge oft
schlimmer“.33
Eine eindeutige Kontrollfunktion übt es jedoch, was wir in dieser Arbeit aufgezeigt
haben, über andere Männchen aus, es verhindert bzw. unterdrückt Kämpfe und dürfte
hierdurch auch mitverantwortlich sein für die beschriebene Peripheralisierung der
Männchen.34
Unsere Versuchsserie zeigte, dass die Rolle des Alpha-Männchens nicht an ein
bestimmtes Männchen gebunden ist, bei Abwesenheit des momentanen Alpha wird
dessen Position von einem anderen Männchen übernommen, das sich dann auch
gegenüber den übrigen Männchen in der Gruppe durchsetzt und die vormals zu diesen
bestehenden Kontakte reduziert.
Das besondere Verhalten des Alpha-Weibchens haben wir ebenfalls bereits ausführlich
beschrieben (Unterkapitel 14.2.2), vergleichbar nur dem Alpha-Männchen
kontrolliert es alle anderen Gruppenmitglieder. Diese Kontrollfunktion wird in
jeder sich bildenden Gruppe von jeweils einem Weibchen wahrgenommen.
Während der vorliegenden Untersuchung suchte Vanda vornehmlich Kontakt
zum Alpha-Männchen, zum Beta-Weibchen (ihrer Tochter Frieda) und zum
Gamma-Weibchen Mecki, solange diese den dritthöchsten Rang in der
Weibchenhierarchie hielt. Zudem unterhielt Vanda auffällig häufig soziale
Kontakte zu Junge tragenden Weibchen, dies vor allem ca. 30 Tage nach der
Geburt.35
Im Gegensatz zu Nikita waren ihre Interaktionen mit anderen adulten Weibchen nicht
nur einseitig, gerade die von ihr besonders präferierten Weibchen Mecki, Vera, Zita
und Stirni putzten sie mehr oder weniger genauso häufig wie Vanda diese.
Kontaktaufnahmen zu den übrigen adulten Weibchen gingen dagegen stets von ihr aus.
Die übrigen juvenilen und subadulten Mitglieder der Gruppe ignorierte sie unabhängig
vom Geschlecht. Im Vergleich zu den ranghohen Weibchen Frieda und Mecki war
ihr Anteil am agonistischen Verhalten gering, hohes agonistisches Verhalten
zeigte sie nur gegenüber dem Gamma-Weibchen Mecki und dessen Tochter
Calva.36
Wir haben bereits beschrieben, dass die Tochter des Alpha-Weibchens Vanda, das Beta-Weibchen Frieda, bereits im Alter von einem Jahr in der Lage war, die übrige Gruppe (mit Ausnahme des Alpha-Männchens) mühelos zu kontrollieren, dies auch in Abwesenheit der Mutter. Während der Versuche wurde ihre hohe soziale Stellung von allen später geborenen juvenilen Männchen und Weibchen respektiert. Sehen wir einmal von den zu Calva und Flava unterhaltenen sozialen Beziehungen ab, gingen Interaktionen zu anderen Tieren stets von Frieda aus. Sie war als passiver Partner nur attraktiv, als sie selber Alpha-Weibchen war, da wurde sie - wenngleich nicht häufig - geputzt. Ihr agonistisches Verhalten richtete sie vor allem gegen Mecki und, besonders auffällig als sie selber Alpha war, gegen andere adulte Weibchen. Im Gegensatz zu Vanda suchte Frieda den Kontakt zu Junge tragenden Weibchen gleich nach Geburt. Ihre Kontaktaufnahme war stets mit agonistischem Verhalten verbunden, was darauf zurückzuführen ist, dass die jeweiligen Mütter ihr auswichen.37
Adulte Weibchen halten vornehmlich Kontakt zu anderen adulten Weibchen, dabei vor
allem zu rangbenachbarten Weibchen (vgl. auch Unterkapitel 14.1). Dementsprechend
haben Veränderungen in der Rangsituation erheblichen Einfluss auf die sozialen
Beziehungen der Individuen.
Neben dieser auf Rangordnung basierenden grundsätzlichen Präferenz konnte
beobachtet werden, dass Junge tragende Weibchen für Junge tragende Weibchen
besonders attraktiv waren, so nahmen z. B. die Kontakte zwischen Blonda und
anderen adulten Weibchen erst zu, als letztere selber Junge bekamen und diese ca.
15–30 Tage alt waren. Dass Mütter den Kontakt zu Müttern suchen bzw. besonders
attraktiv für andere Gruppenmitglieder sind, wird auch aus dem Freiland
berichtet.
Da die Weibchen durch ihre engen Sozialkontakte zu anderen Weibchen die
Beziehungen zwischen den verschiedenen Clanen regeln, sind sie sicher für die Struktur
einer Gruppe vornehmlich verantwortlich. Adulte Weibchen sind offenbar der zentrale
und stabile Teil der Gruppe. Bei der sozialen Körperpflege in der Gruppe sind adulte
Weibchen dementsprechend nicht nur besonders beteiligt, sondern zudem auch
häufiger aktiver als passiver Partner. Demnach zeigen adulte Weibchen stets mehr
soziale Körperpflege als Männchen. Hingegen beteiligen sie sich am Spiel der Gruppe
kaum.38
Parallel hierzu werden sie attraktiver für das Alpha-Männchen, dabei war auffällig,
dass Nikita auch noch nach erfolgter erfolgreicher Kopulation weiterhin die Nähe der
entsprechenden Weibchen suchte, was durch unsere Daten für Omega, Vera und Stirni
sicher belegt werden kann. Besonders auffällig waren zudem in der Gruppe die engen
Kontakte zwischen adulten Weibchen und ihren Kindern, wobei die Kontakte
von Vanda, Mecki und Stirni und ihren Töchtern (Frieda, Calva, Itta) sogar
die intensivsten waren, die jene zu irgendeinem anderen Tier in der Gruppe
unterhielten. Die engen sozialen Beziehungen zu den Kindern wurden vor der Geburt
bzw. in den ersten Tagen und Wochen nach Geburt eines neuen Jungtieres
reduziert.
Alle juvenilen Tiere, Männchen und Weibchen, unterhielten die meisten Kontakte zu ihren Müttern und darüber hinaus auch zu ihren Geschwistern. Bei den Kontakten zu den Geschwistern war das Geschlecht und das Alter ohne große Bedeutung. Die Verwandtschaft, die nach unseren Daten besondere Bedeutung für weibliche Gruppenmitglieder hat, ist jedoch auch für die soziale Stellung männlicher Jungtiere entscheidend.
Bei den Beziehungen zu den eigenen Müttern wurden geschlechtsspezifische
Unterschiede deutlich, so suchten weibliche Jungtiere häufiger den Kontakt zu ihren
Müttern als männliche und waren sogar der aktivere Teil der Mutter-Kind-Dyade.
Männliche Jungtiere hingegen zeigten nicht so auffällige Tendenzen zu ihren Müttern,
entsprechend war dann auch meist die Mutter der aktivere Partner in der
Mutter-Kind-Beziehung. (Hier bildete nur das jüngste Kind der Gruppe, Omegas
Sohn Paul eine Ausnahme, was zwanglos auf sein Alter zurückgeführt werden
kann.)
Diese Unterschiede korrespondieren den Befunden, dass beim Spielen und sozialen
Explorieren männliche Jungtiere häufiger beteiligt sind als weibliche, weibliche
Jungtiere hingegen sind wie adulte Weibchen attraktiver als Partner beim
Kontaktsitzen und bei der sozialen Körperpflege.
Bei allen juvenilen Männchen war besonders auffällig, dass sie außer den
Kontakten zu ihrer Familie nennenswert häufige positiv-soziale Beziehungen nur
zu anderen juvenilen Männchen unterhielten, dabei präferierten sie jeweils
den Kontakt zu ganz bestimmten anderen Männchen, nämlich den Kontakt
zu juvenilen Männchen, deren Mütter der eigenen Mutter rangbenachbart
waren.39
Juvenile Weibchen haben gleichfalls neben den Kontakten zu ihren Müttern und
Geschwistern vor allem soziale Beziehungen zu anderen juvenilen Weibchen, darüber
hinaus jedoch auch zu adulten Weibchen. Bei den juvenilen Weibchen können wir
ebenfalls Präferenzen für bestimmte andere Individuen beobachten.
Juvenile Weibchen haben somit die gleichen Präferenzkriterien wie juvenile
Männchen, darüber hinaus jedoch zusätzlich Kontakte zu den Müttern der von ihnen
präferierten Juvenilen, das gilt für Calva, Flava, Itta, nicht jedoch im gleichen Maße
für Christa. Es scheint so, als ob juvenile Weibchen bevorzugt Kontakt zu
Töchter-Müttern unterhalten, die rangniedriger sind als die eigenen Mütter. Für
die Wahl der Partner ist wohl die Rangposition der Mutter von höchster
Bedeutung.40
Darüber hinaus sitzen sie jedoch auch häufig mit Söhne-Müttern zusammen, zu denen
ihre eigenen Mütter intensive Sozialkontakte pflegen.
Durchgängig galt für alle juvenilen Weibchen, dass diese alle Weibchen, adulte und
juvenile, bedrohen konnten, die von der eigenen Mutter bedroht wurden, dies
unabhängig vom Alter. So waren Calva, Itta und Christa zu Zeiten, zu denen ihre
Mutter das ranghöchste Weibchen war, jeweils unbestritten zweitrangiges Weibchen.
Gleichfalls verloren sie immer dann ihre Rangposition, wenn die Mutter die
Rangposition verlor.
Bei den juvenilen Männchen war neben dem momentanen Rang der Mutter
offensichtlich auch das Alter der Jungtiere von hoher Bedeutung, so bedrohten die
jüngsten Männchen Protus und Paul in der Regel nur dann andere Tiere, wenn diese
auch von ihren Müttern bedroht wurden (Paul drohte allein Jungfrau (E.S. 2.2,
2.5, 2.6, 2.7, 3.3) und Protus (E.S. 3.2 ff), auch als diese nicht von Omega
bedroht wurden, sicherte sich also diesen beiden gegenüber einen höheren
Rang.)
Ältere juvenile Männchen drohten hingegen offensichtlich unabhängig von ihren
Müttern. Für die Richtung der agonistischen Aktivität war der Introduktionszeitpunkt
von hoher Bedeutung, so drohte Max auch Orbi (E.S. 2.4 und 3.5), Ernst (E.S. 2.8
und 3.6) und Tritus (E.S. 2.10 und 3.5); Orbi: Ernst (E.S. 2.7) und Tritus
(E.S. 2.7); und Berni: Tritus (E.S. 2.8), also jeweils den später introduzierten
Männchen. Ganz im Gegensatz zu ihren Müttern nutzten diese Männchen
also die Chance des Platzvorteils voll aus. Dies gelang ihnen jedoch nicht
gegenüber Berni und Majo, die beide sofort nach Introduktion entsprechend ihrer
früheren Position die Rolle eines Alpha-Männchens übernahmen (Berni (E.S. 2.6
und 4.1 ff), Majo (E.S. 2.8 und 4.4)) und gegenüber anderen Individuen der
Gruppe dominierten (auch gegenüber den „künstlich hochrangigen“ Weibchen).
Während sich Majo in der Situation, in der die drei rangniedrigsten Weibchen
„künstlich ranghoch“ waren und seine Mutter die höchste Stellung innerhalb der
Weibchenhierarchie erreichen konnte, sich mühelos gegenüber den ein Jahr älteren
Berni und Orbi durchsetzte, konnte er dies nicht mehr, als er gemeinsam mit Mecki
und Calva in die Gruppe reintroduziert wurde, in der rangmittlere Weibchen
ranghoch waren. Wohl bedingt durch die Tatsache, dass Berni und Orbi in dieser
Gruppe schon wochenlang Kontrollfunktionen wahrnahmen und unbestritten das
Zusammenleben der Gruppe regulierten (E.S. 4.0 ff), gelang es Majo nur anfänglich
sich durchzusetzen (E.S: 4.4), dann aber verlor er die höchste Position an
Berni und sank auf den 3. Platz in der Männchenhierarchie nach Orbi ab.
Auffällig verlor er jedoch, im Gegensatz zu seiner Mutter, den hohen Rang nicht
völlig.
Es könnte jedoch auch sein, dass Majos Verlust der sozialen Position nicht die Folge
sondern Ursache des Rangabgleitens von Mecki war. Für eine gegenseitige
Wechselwirkung zwischen Rang der Mütter und Rang des Sohnes spricht
auch, dass gerade die Mütter der ältesten juvenilen Männchen (Blonda und
Barbara) auch die höchsten Positionen innerhalb der rangmittleren Weibchen
erreichten.
Es sei noch bemerkt, dass das Bestreben, adulte Weibchen und damit prinzipiell auch
die Gruppe zu kontrollieren Charakteristikum gerade drei- bis vierjähriger
juveniler Männchen sein könnte. So versuchte Orbi bereits nach seiner ersten
Introduktion alle nach seiner Mutter introduzierten Weibchen zu kontrollieren.
Während Majo zu diesem Zeitpunkt, ca. zweieinhalbjährig, vornehmlich nur
juvenile Männchen bedrohte. Bei der dritten Introduktion jedoch (nun ebenfalls
dreijährig) richtete er auch agonistische Interaktionen gegen Weibchen (E.S. 4.4
ff.).
Unsere Untersuchungen zur Struktur der sozialen Beziehungen in einer Macaca fascicularis-Gruppe haben ergeben:
1. Die Struktur einer Gruppe bleibt mehr oder weniger unverändert über Jahre erhalten; Individuen, die kurzfristig aus der Gruppe separiert wurden, nehmen nach erfolgter Reintroduktion ihre alte Stellung in der Gruppe wieder ein.
2. Für die Struktur der Gruppe sind drei verschiedene
Komponenten bestimmend, nämlich a) rollenspezifisches
Verhalten41 , b)
Familienbeziehungen42 und c) der
hierarchische Aufbau der Gruppe43
3. Oestruszyklen und Geburten ändern an der grundsätzlichen Struktur der Gruppe
nichts. Doch war auffällig, dass die Weibchen während eines Oestrus ihre Kontakte zu
anderen Gruppenmitgliedern reduzierten und zu dem Alpha-Männchen intensivierten.
Nach erfolgter Geburt wurden sie attraktiver für andere Gruppenmitglieder, nämlich
für andere Mütter, für das Alpha-Weibchen (vor allem ca. 30 Tage nach dem Werfen)
und für die Tochter des Alpha-Weibchens (gleich nach der Geburt), wobei die letzten
beiden Individuen vornehmlich an den Jungtieren interessiert waren, die sie
regelmässig auch „stahlen“ oder zu „stehlen“ versuchten. Sowohl durch Oestruszyklen
als auch durch Geburten wird demnach zwar die soziale Position des einzelnen
Individuums vorübergehend entscheidend verändert, nicht jedoch sein grundsätzliches
Beziehungsgefüge zur Gesamtgruppe.
Die zum Ende der abgeschlossenen Versuchsserie zur Sozialstruktur und zur Rangordnung aus 37 Individuen bestehende Gruppe nahm durch Geburten um neun Tiere zu (Micha bis Lotte, vgl. Tabelle 14.6).44
Während der ersten sechs Lebensmonate wurden die acht 1980 geborenen Kinder täglich 15 Minuten lang beobachtet, wobei jeweils nur ein Kind im Focus des Beobachters stand. Die Daten für jedes Kind sind also unabhängig voneinander erhoben. Die Beobachtungen erfolgten zwischen 15.00 und 17.00 Uhr. Die Reihenfolge der Tiere wurde täglich variiert. Die Interaktionen, die von dem jeweiligen Focus-Tier ausgingen bzw. gegen dieses gerichtet waren, hielten wir in einem Verlaufsprotokoll fest. Protokolliert wurde nach vorgegebenem Katalog.
a) Interaktionen zu anderen Gruppenmitgliedern
Positiv-neutrale Interaktionen45, Positiv aktive Interaktionen46
Agonistische Interaktionen47
b) Interaktionen zwischen Mutter und Kind bzw. ungerichtetes Verhalten des Kindes48
c) Zusätzlich protokollierten die Beobachter nach den 15 Minuten jeweils ihren Gesamteindruck nach folgender Vorgabe: An der Mutter, Neben der Mutter, in der Nähe der Mutter, Weg von der Mutter.
Grundsätzlich wurde jede Interaktion festgehalten. Solche, die länger als eine Minute andauerten, wurden jeweils zu Beginn der darauffolgenden Minute erneut protokolliert.
Von den erhobenen Daten wurden die Summe der positiv-neutralen und der positiv-aktiven Interaktionen, die Zeitangaben „An der Mutter“,„Separiert von der Mutter“ und der protokollierte „Gesamteindruck“ verwandt. Eine weitere Aufsplitterung der Daten erwies sich als nicht sinnvoll49. Zusätzlich zu den Mittelwerten der Interaktionen errechneten wir hier auch den jeweiligen Prozentanteil an den Gesamtinteraktionen eines Monates aus und verglichen diesen mit dem „Erwartungswert“.50
Kontakte zur Mutter
Der enge Kontakt zwischen Mutter und Kind, der während des ersten Lebensmonates
fast die ganze Zeit andauert, nimmt bis zum Ende des sechsten Lebensmonates
kontinuierlich ab. Entsprechend nimmt die Zeit „Ohne Körperkontakt zur
Mutter“ kontinuierlich zu. Bei diesen grundsätzlich gleichen Befunden für alle acht
untersuchten Individuen sind darüber hinaus geschlechtsspezifische und individuelle
Unterschiede auffällig. So werden männliche Kinder schneller selbständig als
weibliche. Individuelle Unterschiede sind erheblich, besonders auffällig bei
Felix und Olga, deren Entwicklung anfänglich retardiert erscheint. Sehen wir
uns hier die Daten genauer an, so erkennen wir, dass dies vor allem darauf
zurückzuführen ist, dass Felix und Olga viel häufiger in der Nähe der Mütter
verblieben als die übrigen Individuen. Hierfür ist sicherlich ein Verhalten der
entsprechenden Mütter (Stirni und Barbara) verantwortlich. Diese hielten
nämlich ihre Jungen bei deren Erkundungsversuchen während der ersten
Lebensmonate am Schwanz fest, so deren Bestreben, sich von der Mutter zu entfernen,
hindernd.
Es sei hier bemerkt, dass die zunehmende Selbständigkeit der Kinder eine aktive Leistung
der Kinder und nicht der Mütter ist. Die Mütter blieben ihren Jungen gegenüber bis
zum Ende des Untersuchungszeitraumes (und auch darüber hinaus) tolerant;
mütterliche Abwehr u. ä. waren für die zunehmende Selbständigkeit der Jungen ohne
Bedeutung.51
Der Vergleich der „Gesamteindruck“-Kurven mit den Zeitkurven ergab zudem, dass
alle Kinder während der ersten Lebensphase bei ihren frühen Erkundungen „Neben
der Mutter“ und „In der Nähe der Mutter“ verbleiben.
Kontakte zu anderen Gruppenmitgliedern
Neben den überaus engen Kontakten zu den eigenen Müttern haben die Jungen bereits
während des ersten Lebensmonaten auch solche zu anderen Tieren der Gruppe. Diese
Kontakte nehmen von Monat zu Monat mehr oder weniger kontinuierlich zu (in
Abhängigkeit von dem zunehmenden Selbständigwerden).
Während des ersten Lebensmonates sind die Jungen fast ausschließlich passive
Partner, Kontakte zu anderen Gruppenmitgliedern gehen in der Regel von letzteren
aus. Auffällig sind in dieser ersten Lebensphase die Aufnahmen von Sozialbeziehungen
durch die beiden ranghöchsten Weibchen, nämlich durch das Alpha-Weibchen Vanda
(zu Neo, Felix, Toto, Olga, Tina und Ursula) und seine Tochter, das Beta-Weibchen,
Frieda (gegenüber allen Jungen).
Zudem beobachteten wir Kontaktaufnahmen nun durch die anderen adulten
Weibchen. Zu weiteren Gruppenmitgliedern bestanden kaum Beziehungen.
Mit zunehmendem Alter nehmen dann die Sozialkontakte aller acht untersuchten
Tiere zu Altersgenossen und älteren Juvenilen kontinuierlich zu.
Es ist jedoch schwer zu entscheiden, inwieweit die Häufigkeit der Interaktionen
wirklich ein Zeichen für engere soziale Beziehungen darstellen oder diese nur
zwangsläufiges Ergebnis der zunehmenden sozialen Entwicklung der entsprechenden
Jungtiere sind. Um dies zu klären, sollten die prozentualen Anteile der einzelnen
Tiergruppen (Altersgruppen, Frieda, Vanda, Nikita, adulte Weibchen) an den
Gesamtinteraktionen mit den nach der Anzahl der entsprechenden Tiere zu
erwartenden Werten verglichen werden. Das Alpha-Männchen Nikita unterhielt stets
wenig Kontakte zu den Kindern, diese blieben für ihn (und er für sie) als Sozialpartner
völlig uninteressant. Für das Alpha-Weibchen waren von den männlichen Kindern
Neo, Felix und, eingeschränkt, auch Toto attraktiv, nicht jedoch Micha. Bei
den weiblichen Kindern präferierte Vanda Olga und Ursula vor Tina und
Angela. Es war hierbei auffällig, dass Vanda besonders enge Beziehungen zu
den Kindern rangniedriger Mütter (Neo, Sohn von Jungfrau; Felix, Sohn von
Stirni; Ursula, Tochter von Alba) unterhielt. Über den Erwartungswerten
liegende Sozialkontakte hatte sie über den gesamten Untersuchungszeitraum zu
Neo und Olga, also dem ersten männlichen und dem ersten weiblichen Kind
dieses Jahrganges. Das Beta-Weibchen Frieda suchte den Kontakt zu allen
Kindern bereits während des ersten Lebensmonates. Während des gesamten
ersten halben Lebensjahres unterhielt Frieda besonders enge Kontakte zu
Micha, der für Vanda nicht attraktiv war (s. o.). Wohl auch im Zusammenhang
mit dem Verhalten ihrer Mutter Vanda waren ihre sozialen Beziehungen zu
Neo und Ursula nicht so intensiv wie diejenigen Vandas. Die übrigen adulten
Weibchen suchten nur während des ersten (Micha, Felix, Toto) bzw. zweiten (Neo)
Lebensmonates häufiger den Kontakt zu männlichen Kindern als zu erwarten war,
weibliche Kinder hingegen blieben während der ersten Lebensmonate attraktiv.
Danach lagen aber auch bei diesen (Ausnahme.: Olga) die Werte der adulten
Weibchen unter den Erwartungswerten. Mit den vierjährigen Jungtieren (1976
geboren) interagierten alle acht untersuchten Kinder während der gesamten sechs
Monate weniger als zu erwarten. Das gleiche gilt für dreijährige Jungtiere
(1977 geboren). Zu zweijährigen Jungtieren (1978 geboren) haben nur Tina
im vierten, Ursula im vierten und fünften und Angela im vierten, fünften
und sechsten Lebensmonat relativ viele Kontakte, dabei handelte es sich fast
ausschließlich um solche zu Calva, einer Tochter des Weibchens Mecki. Zu den
einjährigen Juvenilen (1979 geboren) haben nur die drei erstgeborenen männlichen
Kinder und Tina mehr Kontakte als zu erwarten (s. u.). Durchgängig auffällig
waren jedoch für alle acht untersuchten Individuen die engen Beziehungen zu
Gleichaltrigen.
Neben diesen Präferenzen ist bei einer genaueren Analyse der Daten darüber hinaus
zu erkennen, dass alle untersuchten Tiere bei dem Vergleich mit den jeweiligen
Erwartungswerten mehr Kontakte zu Müttern als zu Nichtmüttern unterhalten. Bei
den Interaktionen mit Altersgenossen und älteren Juvenilen bzw. Subadulten sind
zudem Präferenzen zu Individuen des eigenen Geschlechtes bei weiblichen
Kindern durchgängig zu beobachten. Diese haben jeweils mehr Kontakte
zu weiblichen Individuen als nach den Gesamtkontakten zu den einzelnen
Altersgruppen zu erwarten. Für die männlichen Tiere gilt diese Beziehung
nicht.
Daneben haben wir die relative Häufigkeit der Kontakte zu den eigenen Geschwistern
mit denen der jeweiligen Alters- und Geschlechtsgruppe verglichen. Es zeigt sich
hierbei, dass mit einer einzigen Ausnahme (Beziehungen zwischen Olga und Oscar) alle
Kinder mehr Kontakt zu ihren Geschwistern halten als nach der Gesamthäufigkeit zu
erwarten.
Die vorgestellten Ergebnisse könnten sicherlich auch durch die Reihenfolge der Geburten (alle vier männlichen Kinder wurden vor den weiblichen geboren) beeinflusst sein, was wir in der Diskussion berücksichtigen wollen.
Zur Mutter-Kind-Beziehung
Der durchgängig beobachtete enge Mutter-Kind-Kontakt gilt wohl für alle Species des
Genus Macaca. Besonders intensiv ist dieser Kontakt während des ersten
Lebensmonates, hier verlassen die Jungen zwar gelegentlich die Mutter, was wir das
erste Mal zwischen dem 11. und 24. Tag beobachten konnten, doch sind solche
Explorationen nur kurz und beschränken sich auf die unmittelbare Nähe der Mutter.
Die übrige Zeit verbringen die Jungen mehr oder weniger bewegungslos an der Mutter
hängend, wenngleich wir bereits bei einen Tag alten Kindern beobachten konnten, wie
sie ihren Kopf den Raum betretenden Beobachtern zuwandten, als ob sie bereits am
ersten Tag ihre Umgebung wahrnehmen würden. Nach unseren Befunden an
handaufgezogenen Jungtieren sind sie hierzu sicherlich nicht in der Lage. Der
scheinbar entwickelte Geburtszustand des Neonaten täuscht, vielmehr ist der
junge Makake völlig hilflos und auf die permanente Unterstützung der Mutter
angewiesen.
Bei den seltenen Kontakten zu anderen Gruppenmitgliedern ist das Kind stets
passiver Partner. Besonderes Interesse an den Kindern während dieser Lebensphase
zeigten nur die beiden ranghöchsten Weibchen; zur Kontaktaufnahme suchten diese die
Nähe der entsprechenden Mütter, putzten sie, dabei dann vorsichtig auch das Kind
berührend. In solchen Situationen sind die Mütter äußerst vorsichtig und pressen das
Junge an den Körper.
Im zweiten Lebensmonat, die Bewegungen des Kindes sind nun koordiniert, bleibt der
enge Mutter-Kind-Kontakt erhalten, das Junge wird aktiver und verlässt selbständig
die Mutter zu kurzen Erkundungen, wobei es an anderen Gruppenmitgliedern kaum
interessiert ist. Abwehrverhalten konnten wir nicht beobachten, als Grund
der zunehmenden Selbständigkeit ist die Abwehr durch die Mutter somit
auszuschließen.
Wir konnten im zweiten Lebensmonat zudem häufig beobachten, wie die Jungen von
„Tanten“ getragen wurden. Bei den „Tanten“ handelte es sich in dieser Lebensphase
fast ausschließlich um die ranghöchsten Weibchen, die den Müttern die Jungen
„stahlen“, sie herumschleppten und putzten.
Im dritten Lebensmonat begannen alle Kinder, die bisher nur wenig mit
anderen Gruppenmitgliedern interagierten, mit gleichalten und ein Jahr älteren
Kindern bzw. Juvenilen zu spielen. Hierbei wurden Geschlechtsunterschiede
deutlich, weibliche Kinder spielten nur wenig. Das erste Spiel beobachteten wir
jedoch schon bei einem Kind (Toto) am 18. Lebenstag, in der Regel erst in der
vierten Lebenswoche, nur bei Neo erst zum Ende des zweiten Lebensmonates.
Letzter Befund kann darauf zurückgeführt werden, dass bei diesem keine
gleichalten Spielpartner vorhanden waren, wurde er doch als erster geboren. Im
dritten Lebensmonat nimmt auch das generelle lokomotorische Vermögen der
Kinder weiter zu, sie erkunden nun zunehmend auch entferntere Bereiche des
Geheges.
In den darauffolgenden drei Lebensmonaten werden dann die Kontakte zur
Mutter zunehmend durch Kontakte zu anderen Gruppenmitgliedern ersetzt. Der
enge Mutter-Kind-Kontakt bleibt jedoch auch hier (und später) erhalten.
Insofern unterscheidet sich die generelle Entwicklung der heranwachsenden
Javanermakaken nicht von derjenigen anderer Makakenspecies. Eventuelle geringfügige
Unterschiede dürften eher auf Haltung bzw. Befunderhebung zurückzuführen
sein.
Unser auffälliger Befund, dass männliche Kinder früher selbständig werden als
weibliche, wird auch für andere Makakenarten berichtet. Die mögliche Annahme, dass
unsere Daten eventuell darauf zurückzuführen sind, dass unsere Männchen zeitlich
früher geboren wurden, ist nicht hinreichend, da sowohl innerhalb der Männchen als
innerhalb der Weibchen eine Beziehung zum Alter offensichtlich nicht gegeben
ist.
Zu den Kontakten mit anderen Gruppenmitgliedern
Besonders auffällig waren in der ersten Lebensphase die Kontakte zu den beiden
ranghöchsten Weibchen, in der zweiten Lebensphase diejenigen zu gleichalten
Individuen. Das besondere Interesse der ranghöchsten Weibchen ist sicherlich nicht
eine individuelle Eigenschaft dieser Tiere. Grundsätzlich scheinen alle Makaken,
zumindest alle weiblichen Tiere der Gruppe (s. u.) an den Kindern sehr interessiert zu
sein, doch haben nur die beiden ranghöchsten Weibchen auch die Möglichkeit, sich
ohne Gegenwehr den Müttern zu nähern, die Jungen zu berühren und zu „stehlen“ (s.
o.).
Dabei konnten wir auch beobachten, dass diese beiden Weibchen die Jungen den Müttern
direkt vom Körper rissen, meist warteten sie jedoch bis das Junge sich zumindest wenige
Zentimeter von der Mutter entfernt hatte. In solchen Situationen liefen die jeweiligen
Mütter hinter Vanda bzw. Frieda her und warteten auf ihre Chance, das Junge
zurückzuerhalten.52
Die über den Erwartungswerten liegenden Kontakte der Neugeborenen mit den
übrigen adulten Weibchen können zwanglos darauf zurückgeführt werden, dass sie an
ihrer Mutter hängend mehr oder weniger automatisch auch mit den bevorzugten
Kontaktpartnern der Mutter, nämlich adulten Weibchen, interagierten. Da
wiederum Mütter bevorzugt den Kontakt zu Müttern suchen, wäre auch der
verstärkte Kontakt zu anderen jungetragenden Weibchen hinreichend erklärt.
Bedingt durch die Geburtszeitpunkte können wir hier nicht entscheiden, ob
weibliche Kinder generell mehr Kontakt zu adulten Weibchen unterhalten als
männliche. Zu unserem adulten Männchen (Nikita) unterhielten die Kinder kaum
Kontakte. Bei Interaktionen mit älteren Jungtieren werden geschlechtsspezifische
Unterschiede auffällig. An weiblichen Kindern zeigten fast durchgängig nur weibliche
Jungtiere Interesse. Diese unterhielten stets mehr Kontakte zu jenen als für
die entsprechende Altersgruppe zu erwarten. Bei männlichen Kindern sind
die Daten auf den ersten Blick nicht eindeutig. Hier waren männliche und
weibliche Jungtiere in der Regel entsprechend ihrem Anteil an der entsprechenden
Altersgruppe beteiligt (mit einem geringfügig erhöhten Anteil weiblicher
Jungtiere).53
Der Geschlechtsunterschied wird noch auffälliger, wenn wir nur eine Verhaltensweise,
nämlich das Kampfspiel, betrachten. Männliche Kinder zeigten dieses Verhalten
durchgängig häufiger als weibliche, dies obwohl zumindest theoretisch für alle
weiblichen Kinder mehr Spielpartner zur Verfügung standen. Der geringe Anteil der
weiblichen Kinder an den Spielinteraktionen mit männlichen Kindern könnte
hinreichend damit erklärt werden, dass die weiblichen Kinder entweder noch nicht
geboren oder zu jung waren; weiterhin ist auffällig, dass das Interesse der einjährigen
und älteren Männchen am Kampfspiel mit den Kindern mit zunehmender
Individuenzahl nachließ; ein Säugling als Spielpartner war für diese wohl nur
anfänglich besonders attraktiv. Beides reicht aber nicht aus, um die geringe
Spielhäufigkeit mit weiblichen Kindern zu erklären, vielmehr zeigt hier wiederum der
Vergleich der Daten von den mehr oder weniger gleichalten Toto/Olga/Tina/Ursula,
dass für männliche Individuen eine klare Präferenz für Partner des gleichen
Geschlechts besteht. Bei weiblichen einjährigen und älteren Juvenilen sind hingegen
individuelle Präferenzen unabhängig vom Geschlecht auffällig, generell spielen
sie jedoch mit allen Kindern. Es scheint demnach so, dass bei weiblichen
Individuen die Präferenz für ein bestimmtes Geschlecht gar nicht existiert.
Für die Ergebnisse verantwortlich ist also das Wahlverhalten der männlichen
Individuen.
Es muss hier ausdrücklich betont werden, dass eine Beziehung zur Rangordnung der
Mütter während des ersten Lebenshalbjahres nicht existiert.
Zur Bedeutung der Verwandtschaft sei bemerkt, dass wir zwar bei unseren Kindern
gefunden haben, dass alle Kinder mehr Kontakt zu ihren Geschwistern unterhalten als
für die jeweilige Alters und Geschlechtsgruppe zu erwarten, berücksichtigen wir jedoch
die Anzahl der Kontakte insgesamt, so sind die Beziehungen zwischen Olga und Orbi,
Ursula und Max, Angela und Majo, Felix und Itta und zwischen Toto und Berni) für
das jeweilige Kind nahezu ohne Bedeutung. Bei den übrigen Geschwistern
muss darüber hinaus darauf hingewiesen werden, dass die Kontakte zwischen
Ursula und Rudi), Micha und Sascha, Felix und Heinz und zwischen Toto
und Ernst sich nicht in der Häufigkeit von denen zu bestimmten anderen
Individuen der entsprechenden Alters- und Geschlechtsgruppe unterscheiden.
Insofern sind nur die Kontakte zwischen Tina und Flava/Senia bzw. diejenigen
zwischen Angela und Calva/Cornelia auffällig. Offensichtlich sind demnach
verwandtschaftliche Beziehungen in dem ersten halben Lebensjahr nur für die
Beziehungen weiblicher Kinder zu ihren weiblichen Geschwistern für das einzelne Kind
entscheidend.
Wir können somit feststellen, dass von den drei Grundkomponenten der
Makakensozietät, Rolle, Familie und Rang, der Rang (der Mütter) während des
ersten halben Lebensjahres der Kinder keine Bedeutung hat. Die Familie
(sehen wir einmal von den Beziehungen zur eigenen Mutter ab) scheint nur für
weibliche Kinder wichtig zu sein, aber auch nur dann, wenn diese weibliche
Geschwister haben. Rollengemäßes Verhalten hingegen zeigten alle Kinder
gleichermaßen. Bereits während des ersten halben Lebensjahres können wir zwei
verschiedene „Rollen“ unterscheiden, diejenige weiblicher und diejenige männlicher
Kinder.
Unsere Untersuchung an vier männlichen und vier weiblichen Javanermakaken zur postnatalen Entwicklung während des ersten halben Lebensjahres hat ergeben:
1. Der enge Mutter-Kind-Kontakt nimmt bis zum Ende des sechsten Lebensmonates kontinuierlich ab. Hierfür verantwortlich ist das Bestreben der Kinder, sich von den Müttern zu lösen, nicht jedoch Änderungen im Verhalten der Mutter, diese bleiben während der gesamten sechs Monate ihren Jungen gegenüber tolerant.
2. Interaktionen zwischen dem Alpha-Männchen und den heranwachsenden Jungtieren finden kaum statt, besonders enge Beziehungen bestehen jedoch zu dem Alpha-Weibchen und der Tochter des Alpha-Weibchens. Beide sind während der ersten Lebensmonate an den Kindern sehr interessiert, pflegen diese und tragen sie auch herum, dies offensichtlich gegen den Willen der jeweiligen Mütter.
3. Interaktionen mit anderen adulten Weibchen werden vermehrt nur während des ersten Lebensmonates beobachtet, dabei sind offensichtlich andere Mütter stärker an den Kindern interessiert als Nichtmütter.
4. Ab dem dritten Lebensmonat werden die Jungen aktiver, erkunden ihre Umgebung und suchen den Kontakt zu gleichalten und ein Jahr älteren Tieren. Die Beziehungen zu zwei- bis vierjährigen Juvenilen bzw. Subadulten liegen unter den Erwartungswerten.
5. Bereits während des ersten halben Lebensjahres sind Unterschiede zwischen den Geschlechtern auffällig, so werden männliche Kinder schneller selbständig als weibliche. Auch bei den Interaktionen mit älteren Juvenilen und Subadulten sind klare Geschlechtspräferenzen aufzeigbar. So interagieren männliche Gruppenmitglieder fast ausschließlich mit männlichen Kindern, weibliche hingegen mit Kindern beiderlei Geschlechtes. Die Tendenz männlicher Individuen, Kontakte nur mit Kindern des eigenen Geschlechts aufzunehmen, scheint auch schon bei männlichen Kindern vorhanden zu sein.
6. Alle Kinder unterhalten zwar mehr Kontakte zu ihren Geschwistern als für die Individuen der Alters /Geschlechtsgruppe, zu der die jeweiligen Geschwister gehören, zu erwarten, doch sind von diesen Beziehungen nur solche zwischen weiblichen Kindern und deren weiblichen Geschwistern quantitativ auffällig.
7. Eine Beziehung zwischen dem Verhalten des Kindes und dem Rang der jeweiligen Mutter ist während des ersten halben Lebensjahres nicht erkennbar.
Zu Ende des Untersuchungszeitraumes bestand die Gruppe aus 50 Tieren, von denen 38 in Kassel geboren wurden. In der Tabelle 14.6 (Unterkapitel 14.5.1) hatten wir die Daten der 13 1980 und 1981 geborenen Jungtiere angegeben.54
Während des zweiten Lebenshalbjahres wurden die 1980 geborenen Jungtiere an zwei verschiedenen Tagen der Woche jeweils fünfzehn Minuten beobachtet, also wöchentlich eine halbe Stunde. Dabei stand nur ein Tier im Focus des Beobachters; die Daten wurden also unabhängig voneinander erhoben, und zwar für jedes Individuum über sechs Monate. Der Katalog der zu protokollierenden Verhaltensweisen war mit dem für die ersten sechs Lebensmonate verwandten identisch (vgl. Unterkapitel 14.5.1).
Für die Ermittlung der Ergebnisse addierten wir die gesamten von dem jeweiligen Jungtier ausgehenden bzw. gegen es gerichteten positiv-sozialen Interaktionen, in Bezug zu allen Gruppenmitgliedern bzw. zu allen Individuen der entsprechenden Altersgruppe, zu den gleichgeschlechtlichen Individuen der entsprechenden Altersgruppe, und errechneten den Anteil der Interaktionen der verschiedenen Gruppen, der Männchen und Weibchen, der niederrangigen und mittel- bzw. hochrangigen Individuen und der Geschwister.55 Da die Daten für jedes Individuum unabhängig erhoben wurden, sind - bedingt durch die unterschiedlichen Geburtstermine - die verglichenen Daten meist zu verschiedenen Zeitpunkten erhoben worden. Die Selbständigkeitskurven geben den prozentualen Anteil der gesamten Beobachtungszeit des jeweiligen Monats an, an denen die entsprechenden Jungtiere ohne Kontakt zur Mutter frei agierten. Die Kurven der rangmittleren und rangniedrigen Individuen sind Mittelwerte aller Individuen des entsprechenden Rangbereiches. Auf die Darstellung dieser Kurven habe ich hier verzichtet, sie sind über die entsprechende Publikation ([285]) zugänglich.
Kontakte zur Mutter Auch im zweiten Lebenshalbjahr blieb der enge Kontakt zwischen Mutter und Kind erhalten, wenngleich die Kinder erwartungsgemäß nun noch häufiger von der Mutter entfernt waren als zu Ende des ersten Lebenshalbjahres. Im Mittel waren wiederum männliche Individuen selbständiger als weibliche, individuelle Unterschiede jedoch weit auffälliger als geschlechtsbedingte. Ein weibliches Jungtier (Angela) war zum Ende des ersten Lebensjahres am selbständigsten, zwei männliche Jungtiere (Neo, Felix) hingegen hielten sich wieder häufiger an der Mutter auf.
Kontakte zu anderen Gruppenmitgliedern
Hier haben wir erneut für alle Individuen des 1980er Jahrganges
die Sozialbeziehungen zu anderen Gruppenmitgliedern
angegeben.56
Den Kontakten zu den ranghöchsten Individuen kommt nahezu keine Bedeutung
zu. Nur das Jungtier Angela hatte mehr Kontakte zu Alpha-Männchen und
Alpha-Weibchen als nach der Anzahl der Gesamtkontakte zu allen Gruppenmitgliedern
zu erwarten.
Zum Beta-Weibchen hingegen unterhielten nun alle Jungtiere des Jahrganges
Kontakte. Nur bei Neo und Olga liegen sie unter, bei Micha, Toto und Tina über dem
Erwartungswert. Bemerkenswert war, dass Micha mit diesem Weibchen besonders
häufig interagierte, dies deshalb, da Micha bereits im ersten Lebenshalbjahr
durchgängig von Frieda präferiert wurde. Bei den Kontakten zu adulten Weibchen der
Gruppe liegen die Werte der drei früher geborenen Männchen (Neo, Felix, Micha) und
der von Angela unter dem Erwartungswert, die von Toto, Olga, Ursula und Tina über
diesem. Von den untersuchten weiblichen Kindern zeigt also nur Angela weniger
Kontakte zu adulten Weibchen als zu erwarten; berücksichtigen wir jedoch auch den
Anteil des Alpha-Weibchens, betrachten also die Kontakte der Jungtiere zu allen
adulten Weibchen (ohne die eigene Mutter), dann interagieren alle weiblichen
Jungtiere häufiger mit adulten Weibchen als zu erwarten, auffällig häufig jedoch nur
Olga.57
Betrachten wir nun die Kontakte zu den 1981 geborenen, ein Jahr jüngeren Tieren,
dann haben zwangsläufig und erwartungsgemäß die drei im Jahr 1980 früher
geborenen Männchen nur wenig mit diesen jüngeren bzw. richtiger diesem Jungtier
(nur Lotte wurde im Untersuchungszeitraum geboren) interagiert. Bei Toto, Olga,
Tina und Angela hingegen liegt die Zahl der Kontakte deutlich über dem
Erwartungswert. Auffällig ist demnach nur der geringe Anteil Ursulas an den
Interaktionen mit später geborenen Jungtieren (s. u.).
Bei den Kontakten zu Jungtieren, die im gleichen Jahr geboren wurden, ist auch im
zweiten Lebenshalbjahr aufzeigbar, dass alle Jungtiere mit ihren Altersgenossen
häufiger interagierten als nach der Anzahl der Kontakte und der Anzahl der Jungtiere
zu erwarten. Häufige Kontakte zu ein Jahr älteren Jungtieren, den 1979 geborenen,
finden wir hingegen nur bei den ältesten drei Männchen. Bei den Kontakten zu den
weiteren - noch älteren - Juvenilen fallen nur solche zu vier Jahre älteren
auf, was allein auf Kontakten zu einem Weibchen (Birgit) beruht. Diese war
an den Jungtieren interessiert, pflegte sie bzw. ließ Kontaktaufnahmen zu.
Dabei ist bemerkenswert, dass zu den Kindern ranghöherer Weibchen (Angela,
Tochter eines ranghohen Weibchens, Toto, Sohn des ranghöchsten rangmittleren
Weibchens) bzw. zu Töchtern rangmittlerer Weibchen (Tina, Olga) keine Kontakte
bestanden.
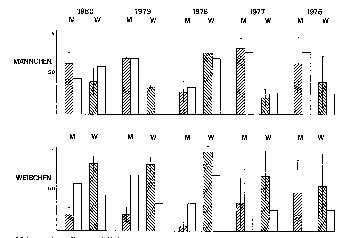
Berücksichtigen wir zusätzlich das Geschlecht der gleichalten und älteren Jungtiere
(Abb. 14.55), dann erkennen wir, dass Männchen mit Juvenilen beiderlei
Geschlechtes, Weibchen hingegen verstärkt nur mit Weibchen interagieren.
Bei den Sozialkontakten weiblicher Juveniler kommt also dem Geschlecht
größte Bedeutung zu. Besonders auffällig ist dieser Befund bei gleichalten
und ein bzw. zwei Jahre älteren Jungtieren. Bei noch älteren Tieren sind
individuelle Präferenzen zu beobachten, bzw. ist Kontaktvermeidung aufzeigbar,
interagieren doch einige subadulte ältere Weibchen überhaupt nicht mit bestimmten
Jungtieren.
Um zu überprüfen, welche Faktoren für dieses Wahlverhalten verantwortlich sein
könnten, haben wir für die einzelnen Jungtiere angegeben, inwieweit sie zu
gleichgeschlechtlichen Jungtieren unterschiedlicher Rangbereiche Kontakte
unterhalten. Echte Wahlmöglichkeiten gab es hier bei den männlichen Juvenilen zu
gleichalten und ein Jahr älteren Männchen, bei den weiblichen Juvenilen zusätzlich
noch zu zwei Jahre älteren Weibchen. Neo, Felix und Ursula suchten häufiger den
Kontakt zu rangniedrigen Individuen, die übrigen fünf Jungtiere hingegen interagierten
häufiger mit rangmittleren oder ranghöheren Tieren. Eine Ausnahme bildet hier nur
Micha bei Kontakten zu Gleichalten (s. u.). Zur Bedeutung verwandtschaftlicher
Beziehungen haben wir schließlich für alle Individuen die Kontakte zu Geschwistern
analysiert. Es zeigten nur Olga und Ursula zu ihren Brüdern weniger Kontakte als
nach der Anzahl der Kontakte zu der Altersgruppe, zu der das Geschwister gehört, zu
erwarten.
Zur Mutter Kind-Beziehung
Im zweiten Lebenshalbjahr bleibt der enge Mutter Kind-Kontakt, der für die ersten
sechs Lebensmonate der acht untersuchten Jungtiere bereits beschrieben wurde
(Unterkapitel 14.5.1), erhalten, nur ändert sich die Qualität der Interaktionen zwischen
Mutter und Kind. Während wir im ersten Lebenshalbjahr nahezu ausschließlich von
der Mutter ausgehende Kontakte beobachteten - die Mutter trägt und putzt das Junge
und bewahrt es vor Gefahr -, beginnt nun das heranwachsende Makakenkind, auch
selbst seine Mutter zu pflegen. Darüber hinaus werden die Kinder, die zwar weiterhin
die Nähe der Mutter suchen, weniger getragen und weniger bewahrt. Kennzeichnend
für das letztere dürfte sein, dass das Festhalten durch die Mutter, wie wir es für
das erste Lebenshalbjahr beschrieben haben, nicht mehr beobachtet werden
konnte. Es muss jedoch offen bleiben, ob hierfür eine Zunahme der Toleranz der
Mutter verantwortlich ist oder aber eine Abnahme des Interesses anderer -
ranghöherer - adulter Weibchen an dem jeweiligen Jungtier. Wir beobachteten
niemals, dass die Jungen von den eigenen Müttern abgewehrt wurden. Wie im
ersten Lebenshalbjahr war demnach die zunehmende Selbständigkeit eine
aktive Leistung des jeweiligen Kindes und keine Funktion des Verhaltens der
Mutter.58
Vernachlässigen wir vorerst das gezeigte Verhalten der Juvenilen und beachten die
soziale Stellung, insbesondere den Rangplatz der Mütter, der bereits früher
umfänglich untersucht wurde (Unterkapitel 14.3.1), dann können wir nach ihrem
Verhalten in neuen sozialen Situationen Angelas Mutter Mecki als „ranghoch“, die
Mütter von Toto (Blonda), Olga (Barbara), Micha (Vera) und Tina (Zita) als
„rangmittel“ und diejenigen von Neo (Jungfrau), Ursula (Alba) und Felix (Stirni) als
„rangniedrig“ klassifizieren, unabhängig von der momentanen sozialen Stellung in der
Gruppe. Tatsächlich entsprach auch im zweiten Lebenshalbjahr der untersuchten
Jungtiere die soziale Stellung der jeweiligen Mutter der nach den Experimenten
gefundenen Rangfolge. Blonda (die Mutter von Toto), die im ersten Lebenshalbjahr
von Angela noch deren Mutter (Mecki) dominierte und die nach Vanda und Frieda
ranghöchstes Weibchen war, verlor wieder diese Position an Mecki, blieb aber
danach weiterhin das ranghöchste Weibchen der übrigen („rangmittleren“ und
„rangniedrigen“) Weibchen.
Untersuchen wir nun, ob eventuell der sozialen Stellung der Mütter für das
Verhalten der Kinder Bedeutung zukommt, und betrachten - getrennt nach
Rangbereichen - die Selbständigkeitskurven von Kindern ranghoher (Angela),
rangmittlerer (Micha, Toto, Tina und Olga) und rangniedriger (Neo, Felix
und Ursula) Weibchen, so war das einzige Kind eines ranghohen Weibchens
selbständiger als alle übrigen sieben Individuen. Darüber hinaus waren Kinder
rangmittlerer Mütter im Mittel selbständiger als diejenigen rangniedriger
Weibchen.
Auffällig ist, dass sich die Kinder der rangniedrigen Weibchen (Neo, Felix und Ursula)
zu Ende des zweiten Lebenshalbjahres besonders häufig an der Mutter oder in der
Nähe der Mutter aufhielten. (Über mögliche Einflüsse des Ranges der Mutter oder aber
der unterschiedlichen Eigenschaften der von ihnen geborenen Kinder für die
Beziehungen zu anderen Gruppenmitgliedern sind damit noch keine Aussagen
gemacht.)
Zu den Kontakten mit anderen Gruppenmitgliedern Für die soziale Stellung des Individuums in der Gruppe sind drei Komponenten verantwortlich, das rollenspezifische Verhalten (Alter und Geschlecht), verwandtschaftliche Beziehungen und der Rangplatz in der Gruppe.
Rollenspezifisches Verhalten Wie bereits für das erste Lebenshalbjahr
beschrieben können wir auch im zweiten Lebenshalbjahr eine unterschiedliche
Orientierung männlicher und weiblicher Kinder aufzeigen. Die Daten erlauben die
Interpretation, dass weibliche Kinder Weibchen präferieren, männliche hingegen kein
ausgeprägtes Wahlverhalten zeigen, entsprechen doch die Anteile älterer weiblicher
Individuen an den Interaktionen männlicher Kinder dem Erwartungswert.
Wie wir noch weiter ausführen werden, ist der Eindruck, dass Wahlverhalten
nur von weiblichen Individuen gezeigt wird, falsch. Verantwortlich für das
scheinbare Fehlen von Geschlechtspräferenzen im zweiten Lebenshalbjahr der
männlichen Kinder ist vielmehr das Verhalten der älteren Juvenilen. Ältere
männliche Individuen sind an weiblichen Kindern nicht interessiert, ältere
weibliche Individuen hingegen zeigen - vor allem - pflegerische Aktivitäten
gegenüber jüngeren Kindern beiderlei Geschlechtes, wie wir es bereits für das erste
Lebenshalbjahr der acht im Focus stehenden Individuen ausführlich beschrieben
haben.
Geschlechtspartnerpräferenzen sind demnach durchgängig aufzeigbar. Entsprechend
interagieren männliche und weibliche Kinder aktiv vor allem mit Individuen des
eigenen Geschlechtes, wie auch bei dem Vergleich der Anteile der Altersgenossen
unterschiedlichen Geschlechtes für männliche und weibliche Individuen überaus
deutlich wird. Hierfür verantwortlich können die geschlechtsspezifischen Unterschiede
im Verhalten sein. Wenn Männchen generell mehr spielen und Weibchen mehr
pflegerische Aktivitäten zeigen, was wir in dieser Arbeit mehrfach beschrieben und
bereits für das erste Lebenshalbjahr aufgezeigt haben, liegt es nahe, in den
unterschiedlichen Verhaltenspräferenzen die Ursache für das unterschiedliche
Wahlverhalten zu suchen.
Überlagert wird die generelle Orientierung zu Individuen des eigenen Geschlechtes
durch eine Grundtendenz aller Jungtiere, bevorzugt enge Beziehungen zu Individuen
zu unterhalten, die in zeitlicher Nachbarschaft geboren wurden. Insofern verwundert es
auch nicht, dass Toto sowohl mit männlichen als auch mit weiblichen Juvenilen gleich
häufig interagierte. Berücksichtigen wir nämlich Totos Alter - er wurde als jüngstes
Männchen geboren und ist nach seinem Geburtsdatum fast gleich alt wie Olga und
Tina - dann können wir seine häufigen Kontakte zu weiblichen Gleichalten zwanglos
mit dieser Grundtendenz erklären. Er scheint für die drei älteren Männchen Neo,
Micha und Felix nicht so attraktiv zu sein, da diese sich selbst als Spielpartner
genügen.59
Vernachlässigen wir das Kalenderjahr und betrachten vielmehr jeweils die sechs
vorangegangenen und sechs folgenden Monate, dann interagieren dennoch alle
Jungtiere verstärkt mit solchen Individuen, die im selben „Jahr“ geboren wurden.
Durch die gleiche Grundtendenz - die Präferenz für Tiere des gleichen Alters - wird
auch der abweichende Befund von Micha bei der Wahl zwischen rangmittleren und
rangniedrigen Sozialpartnern des eigenen Altersbereiches erklärbar, präferierte er doch
hier den etwas älteren Neo und den etwas jüngeren Felix vor dem später geborenen
Toto.
Geschlechtsspezifisches Verhalten dürfte für das Wahlverhalten der drei ältesten
untersuchten männlichen Kinder (Neo, Micha, Felix) bei den Kontakten zu adulten
Weibchen verantwortlich sein. Diese drei Individuen interagierten deutlich weniger mit
adulten Weibchen als zu erwarten, alle weiblichen Individuen dagegen deutlich mehr
mit adulten Weibchen als zu erwarten. Offensichtlich sind im zweiten Lebenshalbjahr
also schon die Qualitäten der Sozialpartner unterschiedlich. Die bereits früh
vorhandenen Unterschiede im Verhalten zwischen den Geschlechtern können -
generalisieren wir unsere eigenen Befunde zum Mutter-Kind-Verhalten - auf ein
unterschiedliches Verhalten der Mütter gegenüber männlichen und weiblichen
Nachkommen nicht zurückgeführt werden.
Verwandtschaft Neben den durch Alter und Geschlecht, also durch das
rollenspezifische Verhalten, bedingten Kontakten zu anderen Gruppenmitgliedern
kommt solchen zu Verwandten generell hohe Bedeutung zu. Dies gilt für alle
Species des Genus Macaca. Ein wichtiger Indikator für engere Beziehungen
ist zweifellos die soziale Körperpflege, die zwischen Geschwistern weitaus
häufiger zu beobachten ist als zwischen anderen Individuen der entsprechenden
Altersstufen.
Wir können demnach vorerst feststellen, dass das Präferieren von Verwandten vor
Nichtverwandten auch das Ergebnis der vorliegenden Untersuchung ist. Besonders
deutlich wurden diese Präferenzen bei gleichgeschlechtlichen Geschwistern, wie wir es
bei den Kontakten zu jedem einzelnen Jahrgang belegen konnten.
Dieses Ergebnis ist aber nur scheinbar eindeutig und muss noch relativiert
werden.60
Messen wir die absoluten Zahlen der Interaktionen zu den anderen Jahrgängen an den
gesamten Interaktionen der jeweiligen Individuen, müssen diese eigentlich
vernachlässigt werden. Die Kontakte zu den Geschwistern können demnach so wichtig
für die soziale Stellung des Individuums nicht sein, wie es die bloße Betrachtung der
Ergebnisse suggeriert.
Rang Die Analyse der sozialen Interaktionen der im Focus stehenden Jungtiere im
ersten Lebenshalbjahr (Unterkapitel 14.5.1) hatte erbracht, dass dem Rangplatz der
Mutter in der Gruppe keine Bedeutung für die sozialen Beziehungen und
die Entwicklung der heranwachsenden Kinder zukommt. Das in der Gruppe
geborene Makakenkind zeigt vor allem rollenspezifisches Verhalten. Im zweiten
Lebenshalbjahr wird der Einfluss des Ranges der Mutter nun deutlich. Wie
bereits aufgezeigt werden Individuen unterschiedlicher Rangbereiche früher oder
später selbständig, bzw. suchen noch häufiger oder seltener die Nähe der
Mütter.61
Wie bereits erwähnt, zeigten die jüngsten der acht untersuchten Jungtiere bei den
Kontakten zu 1981 geborenen Jungtieren besonders häufig Kontaktaufnahmen, was
problemlos erklärt werden konnte (s. o.). Insofern war der geringe Anteil der
Interaktionen zu Individuen dieses Jahres an den Gesamtinteraktionen des Weibchens
Ursula auffällig. Sie interagierte zudem nur mit einem Kind (Lotte) und schien an den
anderen nicht interessiert zu sein. Berücksichtigen wir aber den Rang von Ursulas
Mutter, dann ist diese rangniedrig, was ebenfalls für Lottes Mutter (Omega) zutrifft.
Weit wahrscheinlicher als ein mangelndes Interesse anzunehmen, ist die Vermutung,
dass Ursula nur mit rangniedrigen Kindern interagieren konnte, bei den übrigen
Kindern handelte es sich nämlich nur um Kinder rangmittlerer Weibchen. Ebenfalls im
Verhalten zu Individuen anderer Altersgruppen konnten wir einen Bezug zur
Rangposition der Mütter aufzeigen, so wählten alle untersuchten Individuen im
zweiten Lebenshalbjahr bei Kontakten zu älteren Juvenilen solche des eigenen
Rangbereiches.62
Nämliches gilt auch für die Kontakte zu dem subadulten Weibchen Birgit, interagierte
es doch nur mit männlichen Individuen, die - wie wir oben bereits ausgeführt haben -
den Kontakt zu ihr auch unabhängig von der Mutter suchten, und mit der einzigen
Tochter eines rangniedrigen Weibchens (Ursula). Das gleiche Phänomen, der geringe
Kontakt rangniedriger subadulter Weibchen zu Kindern ranghöherer Mütter erklärt
auch die bereits beschriebene Kontaktvermeidung. Über den Einfluss der Rangposition
der Mütter auf die positiv-sozialen Kontakte der Juvenilen, die wir für die von uns
untersuchte Species diskutieren konnten, liegen aus der Literatur keine Daten
vor.
Unsere Untersuchungen an vier männlichen und vier weiblichen Javanermakaken zur postnatalen Entwicklung während des zweiten Lebenshalbjahres haben ergeben:
1. Der enge Mutter Kind-Kontakt bleibt im zweiten Lebenshalbjahr erhalten. Die Mütter sind weiterhin tolerant zu ihren Nachkommen, das Entferntsein von der Mutter ist stets eine aktive Leistung des jeweiligen Jungtieres.
2. Wie im ersten halben Lebensjahr unterhalten die Jungtiere kaum Kontakte zu dem Alpha-Männchen. Im Gegensatz zu den Befunden des ersten halben Lebensjahres sind nun auch nicht mehr enge Beziehungen zu den ranghöchsten Weibchen der Gruppe, dem Alpha-Weibchen und dem Beta-Weibchen zu beobachten.63
3. Zu den adulten Weibchen der Gruppe unterhalten im zweiten Lebenshalbjahr alle weiblichen Jungtiere mehr Kontakte als nach der Anzahl der Tiere zu erwarten. Generell interagierten alle Jungtiere häufig mit den adulten Weibchen, mit denen auch ihre Mütter häufig interagierten, bzw. hatten wenig Kontakt zu Weibchen, zu denen auch ihre Mütter wenig Kontakt unterhielten.64
4. Wie im ersten halben Lebensjahr ist wiederum auffällig, dass die männlichen Jungtiere auch häufig mit älteren Männchen interagieren, weibliche Jungtiere hingegen beschränken sich auf Kontakte zu weiblichen Individuen, unabhängig von deren Alter.
5. Alle Jungtiere unterhalten mehr Kontakte zu ihren Geschwistern als für die Individuen der Alters- und Geschlechtsgruppe, zu der das jeweilige Geschwister gehört, zu erwarten ist, jedoch sind nur die Kontakte zwischen weiblichen Jungtieren und deren weiblichen Geschwistern quantitativ auffällig.
6. Eine Beziehung zwischen dem Verhalten des Kindes und dem Rang der jeweiligen Mutter ist während des zweiten Lebenshalbjahres belegbar, so sind Kinder rangniedriger Weibchen weniger selbständig als solche ranghöherer Individuen. Darüber hinaus reduziert sich die Anzahl der engen Sozialpartner. Alle Jungtiere interagieren häufig nur noch mit Individuen des gleichen Rangbereiches, also Kinder rangniedriger Mütter mit rangniedrigen Tieren, Kinder ranghoher/rangmittlerer Individuen mit ranghohen/rangmittleren Tieren. Davon nicht betroffen ist der enge Kontakt zu Individuen des gleichen Alters, also zu Tieren, die direkt vor oder nach dem jeweiligen Jungtier geboren wurden.
Offen ist also, worauf das gezeigte Präferenzverhalten beruht, also inwieweit und wie lange junge Männchen für Weibchen attraktiv sind, und ob die Attraktivität gleichgeschlechtlicher Individuen für alle Altersstufen gilt.65 Zudem wollten wir prüfen, ob die Kontakte zu Verwandten durchgängig, also bei allen Tieren der Gruppe, aufzeigbar und ob diese Kontakte für das Individuum von Bedeutung sind, hatten wir doch gefunden, dass im ersten Lebensjahr die tatsächlich vorhandenen Beziehungen quantitativ nicht bemerkenswert sind. Zur Klärung dieser Fragen analysierten wir erneut die Daten der 25-monatigen Studie zur Rangordnung und zur Sozialstruktur der Gruppe (Unterkapitel 14.3 und 14.4) und werteten nun jedoch für jedes Jungtier die Interaktionen zu anderen Gruppenmitgliedern getrennt aus, dabei nach Lebensjahren differenzierend.66
Als Beispiel für das Wahlverhalten der Juvenilen wollen wir die Daten von zwei Jungtieren, Flava und Tritus, im dritten Lebensjahr betrachten. Wir erkennen (Abb. 14.56), dass das weibliche Jungtier Flava am häufigsten mit seiner Mutter Zita, mit den juvenilen Weibchen Calva (28) und Itta (24), den adulten Weibchen Frieda (13), Vera (03), Stirni (09) und Omega (11) und seiner Schwester Senia (33) interagierte, den Kontakten zu männlichen Individuen kam nahezu keine Bedeutung zu. Entsprechend interagierte Tritus vor allem mit seiner Mutter Vera (03) und den juvenilen Männchen Majo (18), Berni (15), Ernst (22), Orbi(16), Max (19), Paul (25), Benni (17) und Vemo (21). Zudem erkennen wir, dass in der Regel die Tiere als passive Partner nur für solche attraktiv sind, deren Kontakt sie auch selber aktiv suchen.
Unabhängig vom Alter und Geschlecht konnten bei den Interaktionen der Juvenilen
untereinander sowohl agonistische als auch positiv-soziale beobachtet werden.
Agonistische Interaktionen gingen jeweils - bei jeder der untersuchten Dyaden - nur
von einem der beiden Sozialpartner aus, so dass bei zwei beliebig ausgewählten Tieren
das eine stets Aktor, das andere stets Empfänger war. Entsprechend den
bisher an der Kolonie erhobenen Daten konnte die Richtung der agonistischen
Aktivitäten nach den Rängen der jeweiligen Mutter (bei weiblichen und männlichen
Individuen) bzw. dem Alter der Tiere (nur bei männlichen) vorhergesagt
werden.
Scheinbar ist aber Flava hierfür kein Beleg, da sie im dritten Lebensjahr auch mit
Individuen agonistisch interagierte, von denen sie selbst bedroht wurde. Dies ist darauf
zurückzuführen, dass Flavas Mutter Zita während der Experimente zur Rangordnung
mehrfach ihren Rangplatz änderte, Flava war insofern manchmal Aktor, manchmal
passiver Empfänger agonistischer Aktivitäten, dies jedoch stets zu unterschiedlicher
Zeit.
Die agonistischen Interaktionen, deren Richtung damit informative Daten zur
Rangstruktur lieferten, hatten jedoch nahezu keinen Aussagewert bezüglich der
Präferenzen der einzelnen Juvenilen untereinander. Es war vielmehr durchgängig
bei Juvenilen unabhängig vom Alter und Geschlecht festzustellen, dass die
Häufigkeit agonistischer Verhaltensweisen keine Aussage über die Häufigkeit
positiv-sozialer Interaktionen zuließ, d. h. es konnte weder eine positive noch eine
negative Korrelation zwischen agonistischem und positiv-sozialem Verhalten
gefunden werden. Da die grundsätzlichen Ergebnisse zur Rangstruktur unserer
Kolonie bereits vorliegen, können wir hier also die agonistischen Beziehungen
vernachlässigen. Wir wollen uns also bei der weiteren Betrachtung auf die Summe der
positiv-sozialen Kontakte beschränken. Zudem wollen wir individuelle Unterschiede
zurückstellen und vorerst nur prüfen, inwieweit das Geschlecht der Juvenilen
und ihr Alter für die beobachteten Präferenzen verantwortlich sind. Da wir
nach unseren bisherigen Ergebnissen wissen, dass der Verwandtschaft hohe
Bedeutung zukommen könnte, haben wir bei der Auswertung nur die Kontakte
zu solchen Individuen berücksichtigt, die nicht über die Mutter verwandt
waren.
Bei allen untersuchten Altersgruppen - sowohl bei weiblichen als auch bei den
männlichen juvenilen Makaken - zeigte sich nun die Bevorzugung des eigenen
Geschlechtes hinsichtlich der positiv-sozialen Kontakte (Abbildungen 14.57 und
14.58).67
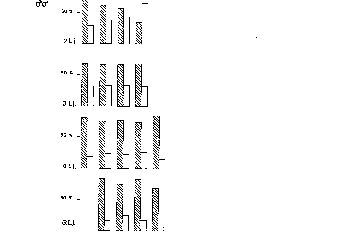
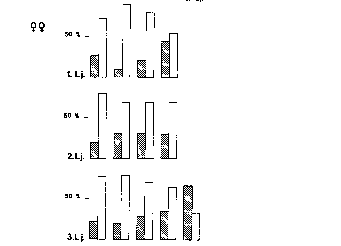
Im Speziellen erkennen wir, dass männliche Jungtiere mehr oder weniger durchgängig
während der gesamten hier ausgewerteten Lebensjahre mehr mit männlichen Juvenilen
interagieren als mit weiblichen. Männliche Individuen unterhalten im zweiten
Lebensjahr hiervon abweichend jedoch relativ mehr Kontakte zu älteren juvenilen
Weibchen als in den späteren Lebensjahren. Dabei scheinen sie mit zunehmendem
Alter der älteren Sozialpartner für männliche Gruppenmitglieder unattraktiver, für
weibliche hingegem attraktiver zu werden, jeweils bezogen auf die Gesamtkontakte zu
allen Individuen der entsprechenden Altersklasse. Besonders bemerkenswert ist
ihre hohe Attraktivität für Weibchen im vierten Lebensjahr, unterhalten sie
doch zu diesen weit mehr Kontakte als zu Männchen der entsprechenden
Altersgruppe.
Weit auffälliger noch als bei den einjährigen Männchen (2. Lebensjahr) wird die
Bevorzugung von Partnern des eigenen Geschlechtes bei älteren männlichen Juvenilen.
Das Alter der Sozialpartner hat hier offensichtlich keinen großen Einfluss mehr.
Durchgängig erkennen wir bei zwei- bis vierjährigen Männchen, dass sie für weibliche
Juvenile unattraktiv sind bzw. auch an diesen kein Interesse haben. Ab dem dritten
Lebensjahr verlieren also männliche Individuen offensichtlich ihre Attraktivität für -
ältere - juvenile Weibchen.
Entsprechend den Befunden an männlichen Individuen finden wir für weibliche
Juvenile, dass diese Individuen des eigenen Geschlechtes vor solchen des anderen
bevorzugen und dies noch weitaus auffälliger als männliche Artgenossen. In der
Abbildung haben wir auch die Ergebnisse für juvenile Weibchen im ersten Lebensjahr
dargestellt, obwohl vergleichbare Befunde bereits für andere juvenile Weibchen
besprochen wurden (Unterkapitel 14.5.1 und 14.5.2). Dies scheint aber wegen der
speziellen Ergebnisse gerade bei weiblichen Kindern geboten, können wir doch nun
diese auf eine breitere Basis stellen.
Ältere weibliche Gruppenmitglieder interagieren, den bisherigen Befunden an der
Kolonie entsprechend, weitaus mehr mit weiblichen Kindern als männliche, was bei ein
Jahr älteren Weibchen besonders deutlich ist. Unabhängig davon, dass die Präferenz
von Individuen des eigenen Geschlechtes - unabhängig vom Alter - erhalten bleibt,
nehmen die Kontakte der juvenilen Weibchen zu männlichen Individuen mit
zunehmendem Alter relativ zu, sie interagieren also relativ häufiger mit männlichen
Juvenilen als im ersten Lebensjahr. Dieser Befund gilt für einjährige und zweijährige
juvenile Weibchen, also für Individuen im zweiten und dritten Lebensjahr. Insofern
überrascht der hohe Anteil der dreijährigen Männchen (4. Lebensjahr) an den
Interaktionen der weiblichen Kinder (1. Lebensjahr) und derjenige der vierjährigen
Männchen (5. Lebensjahr) an den Interaktionen weiblicher Individuen im dritten
Lebensjahr.68
Da bereits bekannt ist, dass alle Juvenilen enge Kontakte zu ihren Geschwistern
halten, haben wir bei der bisherigen Betrachtung der Beziehungen zu anderen
Juvenilen die Interaktionen zu Geschwistern nicht berücksichtigt. Zu Anfang dieses
Kapitels hatten wir bereits exemplarisch gezeigt, dass das Beziehungsgefüge einer
Dyade generell ausgewogen ist, dass also jedes Individuum häufig den Kontakt zu
solchen sucht, die auch selber an ihm interessiert sind. Nämliches gilt für die
Häufigkeit der Interaktionen der Geschwister untereinander. Entweder halten sie also
nur wenige neutrale Kontakte oder aber waren füreinander gleichermaßen
attraktiv und suchten gegenseitig ihre Nähe, dies unabhängig vom Alter.
Über diesen allgemeinen Befund hinaus waren jedoch geschlechtsspezifische
Unterschiede bemerkenswert. Weibliche Juvenile interagierten häufiger mit ihren
Geschwistern als männliche. Zudem fiel auf, dass in jedem einzelnen der Kombination
Schwester/jüngeres Geschwister (unabhängig vom Geschlecht) die ältere Schwester der
aktivere Partner der Geschwisterdyade war. Die Befunde der männlichen Individuen
dagegen waren nicht einheitlich. Der ältere Bruder war also teils der aktivere, teils der
passivere Partner oder aber gleichermaßen oder gleichwenig wie sein jüngeres
Geschwister an diesem interessiert.
Vergleichen wir nun den Anteil der jeweiligen Geschwister an den Gesamtinteraktionen
des Individuums zu der Alters-/Geschlechtsgruppe, zu der das Geschwister gehört,
mit den Anteilen der übrigen Juvenilen - durch die dreizehn in der Gruppe
vorhandenen Geschwisterpaare und die Auswertung nach Lebensjahren haben wir 39
„Geschwisterpaarungen“ analysieren können -, dann war das Geschwister 35ma1 (90
%) attraktivstes Individuum der entsprechenden Alters-/Geschlechtsgruppe, zehnma1
(50 %) zwar nicht am attraktivsten, jedoch attraktiver als nach der Anzahl der Tiere
der entsprechenden Altersgruppe zu erwarten, und nur zweimal (5 %) war der Anteil
des Geschwisters gering. Dem Verwandtsein scheint also durchgängig hohe Bedeutung
zuzukommen.
Neben der Präferenz für Juvenile unterschiedlichen Alters und Geschlechtes haben wir
auch die Interaktionen zu adulten Weibchen erneut überprüft, wobei wir - vorerst - die
jeweiligen Mütter nicht berücksichtigt haben. Es zeigte sich, dass juvenile
Männchen für adulte Weibchen unattraktiv sind. Unabhängig vom Alter der
juvenilen Männchen - wobei hier das erste Lebensjahr nicht berücksichtigt ist -
beschränkten sich die Interaktionen zwischen juvenilem Männchen und adulten
Weibchen auf (geringe) ungerichtete (Kontaktsitzen/Zusammensitzen) und eher
zufällige Kontakte oder aber auf Kontaktaufnahmen durch das jeweilige juvenile
Männchen.
Im deutlichen Gegensatz hierzu kam den juvenilen Weibchen als passive Sozialpartner
adulter Weibchen Bedeutung zu. Besonders häufig wurden jene von solchen adulten
Weibchen gepflegt, die eine ähnliche soziale Stellung wie die eigene Mutter hatten, also
die der Mutter der juvenilen Weibchen rangbenachbart waren. Unabhängig
hiervon waren jedoch die Kontakte zu adulten Weibchen für alle untersuchten
Individuen quantitativ nicht sehr auffällig, weitaus bemerkenswerter dagegen
die Häufigkeit der Interaktionen zur eigenen Mutter. Unabhängig vom Alter
interagierten alle juvenilen Männchen mit ihrer eigenen Mutter weit häufiger
als mit irgendeinem anderen adulten Weibchen der Gruppe. Die Kontakte
zwischen Mutter und Sohn reduzierten sich zwar mit zunehmendem Alter der
Juvenilen, blieben aber selbst im fünften Lebensjahr die intensivsten, die
das jeweilige Männchen zu irgendeinem der adulten Weibchen der Gruppe
unterhielt, wie exemplarisch für Protus und Orbi gezeigt werden soll (Abb.
14.59).
Daneben sei hier schon erwähnt, dass nach neueren Beobachtungen an unserer
Kolonie der enge Kontakt zwischen Mutter und Sohn auch nach dem Adultwerden
erhalten bleibt (Unterkapitel 14.8). Der - wie ich zeigen werde - für männliche Juvenile
erhaltene Befund, dass die Kontakte zur Mutter lebenslang von Bedeutung sind, gilt
gleichfalls für weibliche Individuen.
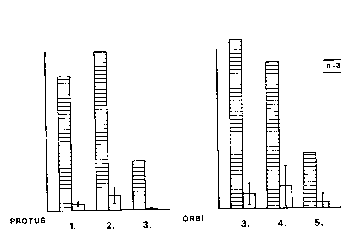
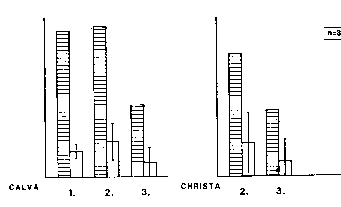
Diese unterhalten zwar generell weit mehr Kontakte zu adulten Weibchen als männliche Juvenile, doch sind diese - messen wir sie an der Zahl der Kontakte zur eigenen Mutter - nur gering, was exemplarisch für Christa und Calva dargestellt sei (Abb. 14.60).
Die Analyse der 25-monatigen täglichen Beobachtungen erbrachte, dass alle in der
Gruppe geborenen Jungtiere - unabhängig vom Alter - den Kontakt zu Tieren des
eigenen Geschlechtes solchem zu Individuen des anderen Geschlechtes vorzogen. Von
diesem generellen Befund abweichend interagierten jedoch einjährige Männchen
(2. Lj.) auffallend häufig und weibliche Kinder (1. Lj.) auffallend wenig mit
dreijährigen juvenilen Weibchen (4. Lj.). Der Befund der männlichen Juvenilen
könnte zwanglos mit dem bereits beschriebenen Verhalten juveniler Weibchen,
pflegerische Aktivitäten gegenüber jüngeren Jungtieren zu zeigen, erklärt werden.
Bei den fraglichen juvenilen Weibchen handelte es sich jeweils nur um ein
bestimmtes Weibchen, Birgit, das - mutterlos - eine rangniedrige Position in der
Gruppe einnahm. Insofern konnten sich ihr alle jungen Männchen nähern,
ohne Gefahr zu laufen, abgewiesen zu werden. Mit dem Tatbestand, dass es
sich hier nur um ein rangniedriges Weibchen - Birgit - handelt, ist auch zu
erklären, dass weibliche Kinder wenig Kontakt zu ihm unterhielten, ließen
doch die Mütter der entsprechenden Kinder Kontaktaufnahmen von und zu
diesem Individuum nicht zu. Der abweichende Befund wäre also hinreichend
erklärt. Die somit stets zu findenden Präferenzen zu Individuen des eigenen
Geschlechtes scheinen jedoch den Ergebnissen an einjährigen Juvenilen zu
widersprechen, hatten wir hier doch gefunden, dass im ersten Lebensjahr Männchen
für Individuen beiderlei Geschlechtes attraktiv sind. Zur Klärung wollen wir erneut
exemplarisch Flava und Tritus betrachten und jeweils die Kontakte zu den übrigen
Gruppenmitgliedern für deren zweites und drittes Lebensjahr angeben (Abb
ma13).
Wir erkennen, dass die Präferenz für Individuen des eigenen Geschlechtes
grundsätzlich vorhanden ist. Vergleichen wir die Kontakte dieser beiden Individuen zu
ein Jahr jüngeren Tieren, dann zeigt sich in beiden Lebensjahren, dass jeweils
Individuen des eigenen Geschlechtes mehr oder weniger deutlich präferiert werden,
darüber hinaus wird aber auch deutlich, dass Flava jeweils häufiger mit ein Jahr
jüngeren Männchen interagierte als Tritus. Tritus - und dies gilt prinzipiell für
alle männlichen Individuen - war mehr an gleichalten und älteren juvenilen
Männchen interessiert, Flava hingegen - wie auch die übrigen juvenilen Weibchen -
interagierte mit jüngeren Weibchen häufiger als mit gleichalten und älteren. Die
Orientierung der Männchen zu älteren und der Weibchen zu jüngeren Individuen ist
demnach für den scheinbaren Widerspruch verantwortlich. Männliche Kinder
interagieren also während ihres ersten Lebensjahres gleichwertig mit älteren
Männchen und Weibchen. Dabei sind sie aber für Männchen immer noch
attraktiver als weibliche Kinder. Insofern ergänzen und spezifizieren diese
Befunde die Ergebnisse an den acht ausgewählten Individuen während des ersten
Lebensjahres.
Neben dieser Orientierung zu Individuen des eigenen Geschlechtes war
erwartungsgemäß der Kontakt zur eigenen Mutter bei allen untersuchten Jungtieren
aufzeigbar. Überraschend war nur, dass selbst bei fünfjährigen Männchen diese enge
Beziehung weiterhin festgestellt werden konnte.
Unabhängig von dieser engen Mutter-Kind-Bindung war auch auffällig, dass alle
Individuen den Kontakt zum jeweiligen Geschwister häufiger suchten als denjenigen zu
Altersgenossen des Geschwisters. Wir müssen aber bedenken, dass nur selten, in
unserem Fall nur bei zwei Geschwisterkombinationen (Berni mit seinem ein
Jahr jüngeren Bruder Ernst, Calva mit ihrer ein Jahr jüngeren Schwester
Cornelia), die Kontakte wirklich für die Individuen quantitativ über denen zu
bestimmten anderen nichtverwandten Sozialpartnern liegen. Der Kontakt zu
Geschwistern war nicht so auffällig wie der zur Mutter. Bedenken wir, dass der enge
Mutter-Kind-Kontakt zwangsläufig bedingt, dass alle Tiere auch mit solchen - zufällig
- mehr interagieren, die zur eigenen Mutter enge Kontakte unterhalten, dann ist der
Kontakt zu Geschwistern durch die Kontakte zur eigenen Mutter hinreichend
erklärt.
Wir können nun also wichten und folgern, dass der Kontakt zu anderen
Gruppenmitgliedern und damit die Struktur der Gruppe vor allem auf rollenspezifischem
Verhalten beruht, alle Individuen halten enge Kontakte zu Tieren des gleichen
Geschlechtes, dabei Individuen des eigenen Altersbereiches, Gleichalte, ein Jahr Ältere
und ein Jahr Jüngere, präferierend. Darüber hinaus unterhalten alle Tiere lebenslang
Kontakte zur eigenen Mutter, wodurch dann zwangsläufig die zu beobachtende enge
Beziehung zu Verwandten, die zweite Komponente, bedingt wird. Überlagert wird
dieses Beziehungssystem schließlich durch die hierarchische Struktur der Gruppe, für
die wir in diesem Teil der Arbeit bei den Beziehungen zu den dreijährigen
Weibchen einen weiteren Beleg aufzeigen konnten. Die Komponenten der
Sozialstruktur der Javanermakaken sind demnach aufgezeigt. Offen ist jedoch, ob und
wieweit Erziehungsprozesse durch andere (ältere) Gruppenmitglieder hierfür
verantwortlich bzw. für die Ausprägung der Komponenten obligatorisch sind. Dies
gilt für alle drei Komponenten der Sozialstruktur. Zur Beantwortung dieser
Fragenkomplexe haben wir Untersuchungen an ohne Einfluss von Müttern und
Geschwistern aufgezogenen Javanermakakenkindern unternommen (Unterkapitel
14.6).
Die Analyse der Interaktionen von 23 in der Gruppe geborenen Jungtieren während einer 25 monatigen Untersuchung zum Sozialverhalten des Javanermakaken hat erbracht:
1. Alle Jungtiere interagieren häufiger mit gleichgeschlechtlichen Individuen aller Altersgruppen als mit solchen des anderen Geschlechtes, dabei präferieren juvenile Männchen gleichalte und ältere Männchen; jüngere Männchen dagegen sind für sie generell unattraktiv, wenngleich sie zu diesen noch mehr Kontakte unterhalten als zu jüngeren Weibchen. Juvenile Weibchen hingegen interagieren mit weiblichen Individuen aller Altersgruppen, für sie sind jüngere Individuen stets attraktiv, besonders solche des eigenen Geschlechtes. Allein auf der Orientierung der Männchen zu gleichalten und älteren und der Weibchen zu jüngeren Individuen beruht der Befund, dass Männchen im ersten Lebensjahr für Individuen beiderlei Geschlechtes gleichermaßen attraktiv sind.
2. Alle Juvenilen unterhalten unabhängig vom Alter und Geschlecht enge Kontakte zur Mutter, die Kontakte zur Mutter bedingen vornehmlich die aufzeigbaren Präferenzen für Geschwister. Bei den Kontakten zu Geschwistern sind ältere Schwestern jeweils der aktivere Partner.
3. Juvenile Männchen sind für adulte Weibchen (Nichtmütter) als Sozialpartner generell unattraktiv. Soweit dennoch Interaktionen zu beobachten sind, ist das jeweilige Männchen der aktivere Partner bei der juveniles Männchen-adultes Weibchen-Dyade.
Juvenile Weibchen dagegen sind als passive Partner für adulte Weibchen attraktiv, besonders für solche, die der Mutter des jeweiligen juvenilen Weibchens rangbenachbart sind.
In vorausgegangenen Unterkapiteln haben wir aufgezeigt, dass drei Komponenten das
Verhalten des Individuums in einer Sozialgruppe des Javanermakaken bedingen, nämlich
1. sein Alter und sein Geschlecht, also sein rollenspezifisches Verhalten, 2. seine
verwandtschaftlichen Beziehungen und 3. seine soziale Stellung im hierarchisch organisierten
Gruppenverband.69
Für alle drei Komponenten wurden zahlreiche Belege gegeben. Mit dem Aufzeigen
dieser Komponenten waren jedoch noch keine Aussagen darüber gemacht, ob und
inwieweit für deren Ausformung Erziehungsprozesse bzw. Einflüsse anderer
(älterer) Gruppenmitglieder verantwortlich sind. Zur Klärung dieser - gerade im
Hinblick auf die kontroverse Diskussion bei unserer eigenen Species - noch
offenen Fragen mussten wir den Einfluss der anderen Gruppenmitglieder auf die
Heranwachsenden ausschließen. Hierzu zogen wir 20 Neugeborene des Javanermakaken
von Geburt an isoliert von der Sozialgruppe auf, so dass diese nur Kontakte zu
Individuen gleicher Aufzuchtsgeschichte und gleichen Alters hatten. Mit diesem
Experimentalansatz sollte es möglich sein, den Einfluss von Lernprozessen für später
zu beobachtenden soziales Verhalten zu bestimmen. Durch unterschiedliche
Experimente und Analysemethoden müsste es gelingen, die Notwendigkeit von
Erziehungsprozessen durch Mütter, Geschwister und andere Gruppenmitglieder
glaubhaft zu machen oder zu widerlegen. Durch Querschnittanalysen sollte die
Komponente „Geschlechtsspezifisches Verhalten“, durch Längsschnittanalysen die
Komponente „Rangordnung“ aufzeigbar sein (vgl. Unterkapitel 14.1). Da zudem
auch einige der Jungtiere eine gemeinsame Mutter haben, müsste auch die
Komponente „Verwandtschaftliche Beziehungen“ analysierbar sein (vgl. Unterkapitel
14.10).
Hier möchten wir über den ersten Teil der Querschnittanalyse, nämlich über das erste
halbe Lebensjahr der Javanermakakenkinder, berichten. Nach den Erkenntnissen
unserer Untersuchung an in der Gruppe aufwachsenden Jungtieren, sollte - soweit
Erziehungsprozesse in der Gruppe hierfür nicht verantwortlich sind - rollenspezifisches
Verhalten schon in dieser Altersphase aufzeigbar sein (vgl. Unterkapitel
14.5.1).70
Vier der für diese Untersuchung verwandten 20 Macaca fascicularis entstammen der Zuchtkolonie der Behringwerke in Marburg (Regina bis Anna), die übrigen 16 sind in Kassel geboren. Die jungen Javanermakaken wurden in zwei Gruppen gehalten, eine (Gruppe 1, Regina - Klara) bestand aus acht (1,7), die andere (Gruppe 2, Armin - Martin) aus zwölf (8,4) Individuen.
Haltungsbedingungen
Die Javanermakakenkinder waren - um eine Beeinflussung durch Artgenossen
auszuschließen - in einem umgebauten Büroraum ohne optischen und akustischen
Kontakt zu anderen Makaken untergebracht. Die Aufzuchtanlage bestand aus einem
vier Kubikmeter großen Käfig, in dem ein zusätzlicher Innenkäfig (1 m x 0,5 m x
0,5 m), angebracht war. Der Innenkäfig diente als Gewöhnungskäfig bei den
Introduktionen der jeweiligen „peers“. Das neu dazukommende Jungtier wurde im
Alter von vier Wochen in den Innenkäfig gesetzt und konnte ohne Verletzungsgefahr
Kontakt zu den anderen - älteren - Artgenossen aufnehmen. Im Alter von fünf Wochen
schließlich wurde das jeweilige Jungtier zu den übrigen „peers“ gelassen. Temperatur
(27 ± 2 Grad C) und Luftfeuchte (60 ± 20 %) waren mehr oder weniger konstant,
das Licht wurde von 8.00 bis 20.00 Uhr geschaltet. Bedingt durch das hohe
Zerstörungspotential der kleinen Makakenkinder und ihre Fähigkeit, jede
Situation zum Verlassen der Käfige zu nutzen, musste der „peer“ - Aufzuchtkäfig
häufig umgestaltet werden (vgl. Unterkapitel 14.1), doch war dies für die hier
beschriebene Lebensphase ohne Bedeutung und kann insofern vernachlässigt
werden.
Beobachtungsbedingungen und -protokoll
Alle Jungtiere wurden ab der fünften Lebenswoche täglich fünfzehn Minuten einzeln
beobachtet, d. h. bei den Beobachtungen stand jeweils nur ein Individuum im Focus
des Beobachters, die Daten für jedes Individuum wurden unabhängig voneinander
erhoben. Beobachtungzeit und -katalog (im Prinzip identisch mit dem in Unterkapitel
14.5.1 angebenen) waren vorgegeben, ebenso die Reihenfolge der Individuen bei den
Beobachtungen.
Für die Auswertung berücksichtigten wir die Daten ab der sechsten Lebenswoche, da
erst hier Interaktionen mit den anderen Gruppenmitgliedern unbegrenzt möglich
waren.
Befundauswertung, Darstellung der Ergebnisse
Von den Daten der 20 Jungtiere haben wir hier (die Befunde der ersten sechs
Lebensmonate) nur diejenigen genutzt, bei denen sichergestellt war, dass die Jungtiere
auch wirklich die Gelegenheit zum Sozialkontakt hatten (Gruppe 1: Anna - Klara,
Gruppe 2: Tobias - Martin). Insofern haben wir die Daten der jeweils ersten Mitglieder
der beiden „peer“ - Gruppen verworfen und für die Auswertung solche von sieben
Männchen und acht Weibchen genutzt. Bei der Auswertung stellte sich heraus, dass
die quantitativen Daten beider Gruppen fast immer im gleichen Bereich lagen,
so dass eine Differenzierung nach Gruppen nur bei einem Verhalten, dem
Kontaktsitzen, nötig war. Grundsätzlich haben wir bei den Abbildungen (der
entsprechenden Publikation ([280]) die Werte für Männchen und Weibchen (nebst
mittlerem Fehler) gesondert angegeben. Wegen des großen Altersunterschiedes der
Gruppenmitglieder in beiden untersuchten „peer“ - Gruppen, war die „Qualität“ der
Sozialpartner unterschiedlich und dementsprechend die Streuung der Werte
groß.
Wir haben daher jeweils zusätzlich innerhalb der Gruppen die Werte von mehr oder
weniger gleichalten Individuen verglichen, sollten diese nicht dem Trend der
Mittelwerte folgen, wird dies gesondert angegeben. Soweit Unterschiede statistisch
eindeutig zu sichern sind, haben wir dies jeweils vermerkt.
Wegen des breiten Datenmaterials und dem Einsatz eines Computerprogrammes war
es möglich, jede einzelne protokollierte Verhaltensweise gesondert zu betrachten,
entsprechend wollen wir hier auch nach Verhaltensweisen getrennt berichten und die
verschiedenen Daten in der Diskussion integrieren.
Betrachten wir das Spielen insgesamt, dann spielen Männchen durchgängig mehr als
Weibchen. Differenzieren wir nach den verschiedenen Spielformen und betrachten
vorerst das sogenannte „neutrale“ Spiel, also das Spiel, wo zwischen aktivem und
passivem Partner nicht unterschieden werden kann, so fällt ein bemerkenswerter
Unterschied zwischen den Geschlechtern auf. Bei dem gemeinsamen Spielen am
gleichen Objekt (ohne klaren Partnerbezug) sind die Weibchen aktiver als
die Männchen, bei dem Kampfspiel und dem Nachlaufspiel hingegen sind
die Männchen die aktiveren. Nur bei dem gemeinsamen partnerbezogenen
Objektspiel ist der Anteil von Männchen und Weibchen ausgewogen. Bei den
gerichteten spielerischen Aktivitäten Spielaufforderung und Spielerisches Beißen
sind die Männchen häufiger aktiver und häufiger passiver Partner als die
Weibchen.
Bei Berücksichtigung von Partnerpräferenzen war auffällig, dass neun der 15 Jungtiere
den Spielkontakt zum nächstälteren Individuum, drei zu dem nächstjüngeren und drei
zu dem nächstälteren des eigenen Geschlechtes suchten. Insgesamt war der
präferierte Spielpartner neunmal gleichgeschlechtlich. Darüber hinaus war
bemerkenswert, dass die jeweiligen Individuen als passive Partner in der Regel
für Männchen attraktiver waren als für Weibchen. Nur in Gruppe 1, in der
nur ein Männchen vorhanden war, wich ein Tier (Klara) von diesem Trend
ab.
Bei dem Kontaktsitzen war es - wie bereits erwähnt - nötig, nach Gruppen zu
differenzieren ([280]). In beiden Gruppen zeigten die Weibchen häufiger dieses
Verhalten als die (Gruppe 2) bzw. das (Gruppe 1) Männchen. Als Partner wurden von
Männchen und Weibchen in der Regel Weibchen gewählt.
Bei dem Verhalten Gehenzu war vor allem auffällig, dass die Jungtiere im ersten
halben Lebensjahr erwartungsgemäß nicht so oft zu anderen Gruppenmitgliedern
gingen, wie sie von diesen aufgesucht wurden.
Bei der Sozialen Körperpflege wird deutlich, dass, wie bei dem Gehenzu, die Tiere
weit häufiger passive als aktive Partner sind, darüber hinaus, dass Weibchen
attraktiver sind als Männchen. Ohne jede Ausnahme war der jeweils aktivste
Sozialpartner älter, meist weit älter, als das gewählte Jungtier.
Als aktive Partner bei dem Manipulieren konnten Unterschiede zwischen
Männchen und Weibchen nicht festgestellt werden, auffällig war, dass elf der
fünfzehn Jungtiere bei dem aktiven Manipulieren ein (noch) jüngeres Tier
präferierten.
Dementsprechend verwundert es nicht, dass der aktivste aktive Partner meist (in vier-
zehn der fünfzehn Fälle) älter war als das jeweilige Jungtier. Zudem wurden eindeutig
Weibchen bevorzugt von anderen Gruppenmitgliedern manipuliert.
Das Verhalten Umarmen wird von Männchen und Weibchen unterschiedlich häufig
gezeigt, Weibchen umarmen andere Gruppenmitglieder - unabhängig vom Alter und
Geschlecht - häufiger als Männchen. Als passive Partner, also als Partner, die
von anderen Gruppenmitgliedern umarmt werden, sind Weibchen weitaus
attraktiver als Männchen. Darüber hinaus ist bemerkenswert, dass alle Individuen,
Männchen und Weibchen, bevorzugt von älteren Gruppenmitgliedern umarmt
werden.
Bei dem Klammern sind die von uns untersuchten 15 „peers“ häufiger aktiver als
passiver Partner. Sie klammerten bevorzugt an älteren (14 der 15 „peers“)
Gruppenmitgliedern. Interessant ist zudem, dass Männchen stets Weibchen,
Weibchen hingegen Individuen beiderlei Geschlechtes zum Klammern bevorzugt
aufsuchten. Als passive Partner waren die weiblichen Kinder attraktiver als die
männlichen.
Neben den bereits beschriebenen Verhaltensweisen haben wir noch betrachtet, wie
häufig unsere 15 Jungtiere von anderen Gruppenmitgliedern getragen wurden,
bzw. wie oft hieran Männchen und Weibchen beteiligt waren. Die gefundenen
Werte des Tragens haben wir mit dem Erwartungswert (EW) verglichen,
also dem Wert, den wir erwarten müssten, wenn Männchen und Weibchen
gleichermaßen am Tragen der Jungen beteiligt sind. Es zeigte sich, dass das
Tragen durch - meist - ältere Gruppenmitglieder eine „Tätigkeit“ der Weibchen
ist.
Die übrigen protokollierten Verhaltensweisen waren quantitativ nicht bemerkenswert.
Erwähnt werden soll jedoch, dass bei dem Verdrängen erwartungsgemäß die
„peers“ weit häufiger passiver als aktiver Partner sind.
Bei den agonistischen Interaktionen schließlich, bei denen die „peers“ im ersten
halben Lebensjahr - ebenfalls erwartungsgemäß - eher die passiveren Partner sind,
zeigten sich dennoch bei den geringen von ihnen ausgehenden Aktionen (Männchen im
Mittel 35, Weibchen im Mittel 13) Unterschiede. Weibchen bedrohten (Drohen mit
und ohne Lautäußerung) und jagten andere Gruppenmitglieder, Männchen
hingegen traktierten und bissen die Sozialpartner. Es sei noch erwähnt, dass
Weibchen im Mittel 102 und Männchen im Mittel 118 agonistische Aktivitäten
„erlitten“.
Wie bereits anfangs betont unterscheiden sich ohne jeden Zweifel männliche und
weibliche Individuen in ihrem Verhalten. Offen ist jedoch, wodurch diese Unterschiede
bedingt sind. Nach Literaturdaten (zitiert in [280]) werden Neugeborene bereits kurz
nach Geburt von anderen Gruppenmitgliedern untersucht und manipuliert,
das Geschlecht - so die Annahme - wird bestimmt. Zahlreiche vergleichbare
Beobachtungen konnten wir ebenfalls an unseren Kolonien des Javanermakaken und
des Kapuzineraffen machen. Die Beobachtungen, dass Mütter männliche Kinder früher
und häufiger bedrohen und bestrafen, suggerierten, dass das Verhalten der Infantes
und Juvenilen eine Funktion des Verhaltens ihrer Umgebung ist. Weit eher als durch
direkte Beobachtungen ließe sich das Geschlecht des Jungtieres nach dem
Verhalten der Mutter bestimmen. Demnach spräche alles dafür, dass zu Beginn
Geschlechtsunterschiede gering sind. Literaturdaten stützten die Hypothese, dass
die geschlechtsspezifischen Unterschiede im Verhalten vor allem durch die
Behandlung durch Artgenossen bedingt seien. Solche Befunde stützten zweifellos die
gängigen Annahmen der westlichen Kulturgesellschaft, geschlechtsspezifische
Unterschiede wären demnach vor allem erziehungsbedingt. Unsere eindeutigen
Befunde an in der Gruppe aufgezogenen Jungtieren bzw. auch bereits bekannte
Unterschiede im Verhalten der Geschlechter können diese Annahme nicht
widerlegen.
Wir haben daher nach einer Möglichkeit gesucht, den Einfluss der sozialen Umwelt zu
relativieren und haben dies durch Handaufzucht vom ersten Lebenstag an
erreicht. Bei den untersuchten „peers“ können wir sicher sein, dass sie Einflüssen
adulter Individuen nicht unterworfen waren. Sollten die „peers“ selber einen
„prägenden“ Einfluß gehabt haben, müssten Unterschiede nicht zwischen den
Geschlechtern, sondern zwischen den Gruppen auftreten, in einer waren nämlich die
ältesten Individuen weiblich (Gruppe 1), in der anderen männlich (Gruppe 2). Für
solche Einflüsse gab es jedoch keinen Beleg.
Der einzige Unterschied, der zwischen den Gruppen auftrat, war die Häufigkeit des
Kontaktsitzens, hier zeigten alle Individuen der Gruppe 2 dieses Verhalten häufiger als
solche der Gruppe 1, was wir bei der Darstellung der Ergebnisse berücksichtigt
haben.
Betrachten wir nun diese Befunde im Einzelnen, so zeigte sich - in Übereinstimmung
mit unserem bisherigen Wissensstand -, dass Weibchen mehr Kontaktsitzen zeigen als
Männchen und Männchen mehr spielen als Weibchen bzw. gleichfalls als Spielpartner
attraktiver als Weibchen sind. Die Differenzierung nach verschiedenen Spielformen
erbrachte darüber hinaus, dass dies für Kampfspiel, Nachlaufspiel, Spielerisches
Beißen und für die Häufigkeit der Spielaufforderung gilt, bei dem Objektspiel
hingegen zeigten sich solche Unterschiede nicht, bei dem Gemeinsamen Spiel
am gleichen Objekt waren die Weibchen sogar aktiver als die Männchen.
Betrachten wir die Resultate von in der Gruppe aufgezogenen Jungtieren im ersten
Lebenshalbjahr bzw. generell die Befunde der (älteren) Juvenilen, dann kann die dort
gefundene höhere Beteiligung männlicher Individuen am Spiel sicherlich durch
Umwelteinflüsse nicht hinlänglich erklärt werden. Männchen spielen offensichtlich
mehr, weil sie Männchen sind, und nicht, weil sie wie Männchen behandelt
werden.
Damit haben wir jedoch die Ergebnisse an den „peers“ nicht ausgeschöpft, neben den
an sich erwarteten Befunden, dass alle „peers“ signifikant häufiger passive
Partner bei dem Verhalten Gehenzu und bei der sozialen Körperpflege bzw.
signifikant häufiger aktiver Partner bei dem Klammern an anderen Tieren sind,
haben wir Geschlechtsunterschiede gefunden, die bisher nicht beschrieben
sind.
Die weiblichen „peers“ waren nämlich bei der sozialen Körperpflege, bei dem
Manipulieren und bei dem Umarmen als passive Partner signifikant attraktiver
als männliche Artgenossen. Da alle „peers“ wegen des Alters der anderen
Gruppenmitglieder zu bestimmten Verhaltensweisen nicht gezwungen werden
konnten, ist der Schluß zwingend, dass weibliche Individuen sich lieber putzen,
manipulieren und umarmen lassen als männliche Individuen, dies bereits im
ersten Lebenshalbjahr. Diesen Befunden korrespondiert, dass die untersuchten
„peers“ signifikant häufiger von weiblichen (älteren) Gruppenmitgliedern getragen
wurden als von (älteren) Männchen. Bei dem Klammern, dem Aufsuchen
meist älterer Individuen mit dem Ziel des engen Körperkontaktes, präferierten
männliche Individuen stets Weibchen, insofern verwunderte es nicht, dass die hier
untersuchten weiblichen „peers“ bereits im ersten Lebenshalbjahr als passive
Partner bei dem Klammern signifikant attraktiver waren als ihre männlichen
Altersgenossen.
Die Unterschiede zwischen den Geschlechtern sind also durchgängig, weibliche
Individuen sind eher Empfänger pflegerischer Aktivitäten als männliche Individuen,
Männchen spielen mehr. Ohne Zweifel ist dieser Geschlechtsunterschied bereits früh in
der Verhaltensontogenese angelegt, einen Einfluss der Umwelt erfordert er, zumindest
bei dem Javanermakaken, nicht.
Die Untersuchungen an ohne Kontakt zu Müttern, Geschwistern und älteren Artgenossen aufgezogenen Javanermakakenkindern haben für das erste halbe Lebensjahr erbracht:
1. Weibliche Jungtiere zeigen häufiger Kontaktsitzen, männliche häufiger Spielen. Bei einer Differenzierung zwischen verschiedenen Spielformen zeigt sich, dass Männchen bei dem Kampfspiel, bei dem Nachlaufspiel, bei dem Spielerischen Beißen und bei der Spielaufforderung aktiver sind als Weibchen. Weibchen dagegen beteiligen sich häufiger an dem Gemeinsamen Spiel am gleichem Objekt.
2. Männchen und Weibchen sind bei dem Verhalten Gehenzu und bei der Sozialen Körperpflege signifikant häufiger passive und bei dem Klammern signifikant häufiger aktive Partner, d. h. sie werden von anderen (älteren) Gruppenmitgliedern häufiger aufgesucht und nutzen diese zum Klammern.
3. Weibchen sind bei der Sozialen Körperpflege, bei dem Manipulieren und bei dem Umarmen als passive Partner signifikant attraktiver als Männchen.
4. Männchen und Weibchen werden von (älteren) weiblichen Gruppenmitgliedern weitaus häufiger getragen als von männlichen. Diesem Befund korrespondiert, dass die untersuchten Weibchen bei dem Klammern als passive Partner signifikant attraktiver waren als männliche Artgenossen.
Durch die Analyse der Daten des zweiten Lebenshalbjahres wollen wir prüfen, ob die bereits im ersten Lebenshalbjahr aufgezeigten noch geringen Unterschiede zwischen den Geschlechtern auch im zweiten Lebenshalbjahr vorhanden sind, bzw. ob diese und weitere Unterschiede während dieser Lebensphase reifen.71 Darüber hinaus prüfen wir, inwieweit dem Alter und dem Geschlechts der „peers“ bei der Wahl der Sozialpartner Bedeutung zukommt.
Das Vorgehen habe ich in Unterkapitel 14.6.1 angegeben. Die zu analysierenden Daten beruhen hier auf Beobachtungen aller zwanzig Jungtiere, auf Einzeltier-Beobachtungen (täglich 15 Minuten) von neun Männchen und elf Weibchen.
Offensichtlich altersbedingt nimmt die Häufigkeit sozialer Interaktionen während
des zweiten Lebenshalbjahres auffällig zu. Diese Zunahme lässt sich für die
verschiedenen Spielformen gut sichern (bei männlichen Individuen nimmt die mittlere
Häufigkeit des Kampfspieles sogar um mehr als 300 % zu). Darüber hinaus ist
besonders auffällig, dass Männchen und Weibchen im zweiten Lebenshalbjahr
andere Individuen weitaus häufiger aktiv putzen (Zunahme bei Weibchen
um über 700 %) und häufiger geputzt werden (nur Weibchen) als im ersten
Lebenshalbjahr. Insofern ist ebenfalls bemerkenswert, dass die Jungtiere in dem hier
besprochenen Lebensabschnitt weniger häufig umarmt, manipuliert (mit den Händen
angefasst) und umklammert werden als für die ersten sechs Lebensmonate
berichtet.
Bei dem „neutralen“ Spiel, also dem Spiel, bei dem nicht nach aktiven und passiven
Partnern getrennt werden kann, ist auffällig, dass Männchen häufiger das Verhalten
Kampfspiel zeigen als weibliche Individuen, diese spielen jedoch häufiger mit Objekten
als ihre männlichen Altersgenossen. Unterscheiden wir zwischen partnerbezogenem
Objektspiel und Objektspiel ohne klaren Partnerbezug lässt sich für letzteres die
höhere Beteiligung weiblicher Individuen sichern. Berücksichtigen wir zusätzlich noch
das Nachlaufspiel, wird auch hier eine höhere Spielaktivität männlicher Individuen
auffällig. Bei dem gerichteten Spielen schließlich fällt auf, dass Individuen beiderlei
Geschlechtes zwar etwa gleich häufig andere Gruppenmitglieder zum Spielen
auffordern bzw. von diesen aufgefordert werden, dass aber Männchen bei dem
spielerischen Beißen sowohl als aktive als auch als passive Partner häufiger beteiligt
sind.
Bei dem Kontaktsitzen lassen sich keine geschlechtstypischen Unterschiede aufzeigen
(s. u.).
Bei dem Gehenzu hingegen ist auffällig, dass Männchen und Weibchen etwa gleich
häufig von anderen Individuen aufgesucht werden, dass aber männliche Individuen in
dieser Lebensphase häufiger andere Gruppenmitglieder aufsuchen als weibliche
Individuen.
Betrachten wir die Beteiligung beider Geschlechter an der sozialen Körperpflege, wird
eine höhere Aktivität der Weibchen offensichtlich, diese putzen andere Individuen
häufiger bzw. werden häufiger geputzt. Entsprechendes gilt für das Manipulieren
und das Umarmen, wobei sich der Unterschied zwischen den Geschlechtern
jedoch nur für das passive Manipulieren und das aktive Umarmen sichern
lässt. Bei dem Klammern sind Weibchen als passive Partner attraktiver als
Männchen.
Überprüfen wir nun das Wahlverhalten der Gruppenmitglieder, so wird deutlich, dass
alle Individuen Sozialpartner bevorzugen, die vor oder nach ihnen geboren wurden.
Vergleichen wir die Befunde der Männchen und Weibchen, so ist bei dem
„neutralen“ Spiel bemerkenswert, dass grundsätzlich Sozialpartner des eigenen
Altersbereiches unabhängig vom Geschlecht gewählt werden. Soweit (minimale)
Unterschiede bei dem Wahlverhalten beobachtet werden, ist auffällig, dass
nahezu durchgängig männliche Individuen häufiger mit „Nächstälteren“ und
„Nächstjüngeren“ des eigenen Geschlechtes spielen als mit dem „Nächstälteren“ und
„Nächstjüngeren“ unabhängig vom Geschlecht des Partners; für Weibchen dagegen
finden wir durchgängig entgegengesetzte Befunde, d. h. für Individuen beiderlei
Geschlechtes sind nächstältere und nächstjüngere Männchen als Spielpartner
attraktiver.
Die höhere Attraktivität männlicher Sozialpartner für weibliche Individuen ist bei der
Spielaufforderung (aktiv und passiv) und bei dem spielerischen Beißen (aktiv und
passiv) nun ohne Ausnahme zu beobachten. Das gleiche gilt grundsätzlich für
männliche Individuen. Dabei lässt sich allerdings nur die höhere Attraktivität der
Männchen bei dem spielerischen Beißen für nächstältere männliche Individuen sichern.
Darüber hinaus ist auffällig, dass weibliche Individuen aktiv bei dem spielerischen
Beißen nächstjüngere Individuen bevorzugen, bzw. dementsprechend häufiger von
nächstälteren als passive Partner gewählt werden. Bei dem Kontaktsitzen ist
eine Vorliebe für Individuen des eigenen Altersbereiches nicht so auffällig.
Bemerkenswert erscheint jedoch, dass männliche Individuen häufiger mit
nächstjüngeren (unabhängig vom Geschlecht) in engem Körperkontakt sitzen als mit
Nächstälteren und Nächstjüngeren des eigenen Geschlechtes, d. h. Männchen
bevorzugen bei dem Kontaktsitzen nächstjüngere Weibchen. Bei dem aktiven und
passiven Gehenzu kommt dem Geschlecht der Sozialpartner keine Bedeutung zu.
Bemerkenswert erscheint jedoch bei den untersuchten Weibchen der im Vergleich zu
den Männchen höhere Anteil von Individuen des eigenen Altersbereiches.
Bei den übrigen hier besprochenen Verhaltensweisen (Soziale Körperpflege, Manipulieren,
Umarmen, Klammern) finden wir keine Bevorzugung von Individuen des eigenen
Altersbereiches.72
Wir haben bereits erwähnt, dass die sozialen Interaktionen der Javanermakaken im
zweiten Lebenshalbjahr zunehmen (s. o.), was vor allem auf die altersbedingte höhere
Mobilität der Jungtiere zurückgeführt werden kann. Wie im ersten Lebenshalbjahr
spielen Männchen häufiger als Weibchen mit anderen Sozialpartnern. Im zweiten
Lebenshalbjahr kann dieser Unterschied für das Kampfspiel und das spielerische
Beißen gesichert werden. Darüber hinaus konnte erneut beobachtet und nun gesichert
werden, dass weibliche Individuen häufiger an Objekten spielen als männliche.
Unterscheiden wir zwischen partnerbezogenem Objektspiel und solchem ohne klaren
Partnerbezug, dann ist dieser Unterschied - wie im ersten Lebenshalbjahr - nur für die
zweite Form des Objektspieles zu belegen.
Bei dem Gehenzu sind die jungen „peers“ nun ebenso häufig aktive wie passive
Partner, was wiederum ihrer altersbedingten höheren Mobilität entspricht.
Entsprechendes gilt für die soziale Körperpflege; bei dem Manipulieren und Umarmen
hingegen sind sie nun sogar häufiger aktive als passive Partner (s. u.). Bereits im
ersten Lebenshalbjahr suchten Männchen häufiger andere Gruppenmitglieder auf als
Weibchen, im zweiten Lebenshalbjahr wird dieser Unterschied zwischen den
Geschlechtern sicherbar. Entsprechend den Befunden im ersten Lebenshalbjahr sind
Weibchen dagegen bei der sozialen Körperpflege sowohl als aktive als auch als passive
Partner aktiver bzw. attraktiver als männliche Individuen. Waren die Weibchen im
ersten Lebenshalbjahr vor allem Empfänger pflegerischer Aktivitäten, so sind
sie nun auch aktiv am Pflegen anderer Individuen beteiligt. Entsprechende
geschlechtstypische Unterschiede konnten ebenfalls für das aktive Umarmen und
Manipulieren gezeigt werden, was sich für das Umarmen sichern ließ. Den
Befunden des ersten Lebenshalbjahres entsprechend sind weibliche Individuen bei
dem Klammern als passive Partner attraktiver als männliche Artgenossen,
doch geht die Häufigkeit dieser Verhaltensweise - ebenfalls altersbedingt -
zurück.
Im Gegensatz zu den Befunden des ersten halben Lebensjahres konnte bei dem
Zusammensitzen in engem Körperkontakt kein Unterschied zwischen den
Geschlechtern aufgezeigt werden, Männchen und Weibchen zeigen dieses Verhalten im
Mittel etwa gleichhäufig. In Unterkapitel 14.6.3 werden wir überprüfen können, ob die
für Makaken generell beschriebene höhere Beteiligung der Weibchen an diesem
Verhalten bei den handaufgezogenen „peers“ noch „reift“, oder aber, ob der höhere
Anteil der weiblichen Individuen vor allem durch das Präferenzverhalten älterer
(adulter) Weibchen bedingt ist.
Überaus auffällig war, dass alle Individuen bevorzugt mit Tieren des eigenen
Altersbereiches Kontakte unterhielten, wobei eine Tendenz erkennbar (aber nicht
sicherbar) ist, beim Spielen männliche und beim Pflegen weibliche Individuen zu
bevorzugen. Zur Gewichtung dieses Befundes wollen wir Ergebnisse von in der Gruppe
aufgezogenen Jungtieren heranziehen. Wir hatten belegt (s. o.), dass alle in der
Gruppe geborenen Jungtiere Individuen des eigenen Alters bevorzugen. Dieser
„peer-group“ Effekt kann für das hier gefundene Wahlverhalten nur bedingt
verantwortlich sein, sind doch die Individuen der jeweiligen „peer“-Gruppe alle
Individuen des gleichen Altersbereiches. Zudem hatten wir jedoch herausgefunden,
dass alle Individuen bevorzugt mit solchen des eigenen Rangbereiches Kontakte
unterhalten. Bei den hier besprochenen „peers“ war die Rangordnung abhängig vom
Alter. Ohne Ausnahme waren ältere Tiere ranghöher, dementsprechend der
jeweils nächstältere bzw. nächstjüngere Sozialpartner auch der Partner des
eigenen Rangbereiches. Zudem haben unsere Beobachtungen bei der Bildung der
Ausgangsgruppe unserer Kolonie belegt, dass anfänglich der Rangplatz abhängig vom
Zeitpunkt des Einfügens in die Gruppe ist, später hinzugekommene Tiere sind
rangniedriger als früher hinzugekommene. Dafür erbrachten auch Experimente zur
Änderung der Sozialstruktur zahlreiche Belege (s. o.). Bei den hier untersuchten
„peers“ entspricht nun aber die Reihenfolge des Hinzukommens dem Alter. Insofern
können wir hier noch nicht abschließend werten, ob der Rangplatz oder das Alter für
das beobachtete Wahlverhalten verantwortlich ist. Dies blieb weiteren Analysen für
Zeiten, wo der Rang der „peers“ nicht mehr altersabhängig ist, vorbehalten
(s. o.). Unabhängig von dieser Analyse besteht jedoch das Ergebnis, dass
bei nahezu allen beobachteten Verhaltensweisen den Individuen des eigenen
Altersbereiches hohe Bedeutung zukommt. Nur für einige, quantitativ weniger
auffällige, pflegerische Aktivitäten (Soziale Körperpflege, Manipulieren, Umarmen,
Klammern) werden die Sozialpartner nach anderen Kriterien gewählt, hier sind als
passive Partner (Soziale Körperpflege, Manipulieren, Umarmen) bzw. als aktive
Partner (Klammern) jüngere Individuen besonders beteiligt, und umgekehrt. Dieser
Befund bestätigt frühere Ergebnisse, nach denen weibliche Individuen bevorzugt
jüngere Individuen - unabhängig vom Geschlecht - pflegen und tragen. Offen bleibt
somit nur, ob im weiteren Lebensablauf weitere geschlechtstypische Unterschiede
reifen.
Die Untersuchungen an ohne Kontakt zu Müttern, Geschwistern und älteren Artgenossen in einer Spielgruppe („peer-group“) aufgezogenen Javanermakakenkindern haben für das zweite Lebenshalbjahr erbracht:
1. Männliche Individuen spielen häufiger mit anderen Sozialpartnern als weibliche. Dieser Unterschied ist bei dem Kampfspiel und dem spielerischen Beißen sicherbar. Weibchen hingegen spielen signifikant häufiger mit Objekten ohne klaren Partnerbezug.
2. Männliche Individuen gehen häufiger aktiv zu anderen Gruppenmitgliedern, weibliche Individuen hingegen zeigen häufiger pflegerische Aktivitäten bzw. werden auch häufiger von anderen Gruppenmitgliedern hierfür als passive Partner gewählt.
3. Bei der Wahl der Sozialpartner kommt dem Alter der Partner hohe Bedeutung zu. Grundsätzlich werden diejenigen Individuen bevorzugt, die vor bzw. nach dem jeweils beobachteten Tier geboren und der Gruppe hinzugefügt wurden. Darüber hinaus ist im zweiten Lebenshalbjahr eine Tendenz zu erkennen, für spielerische Aktivitäten männliche und für pflegerische Aktivitäten weibliche Sozialpartner zu bevorzugen. Insgesamt gesehen kommt in dieser Lebensphase dem Geschlecht der Sozialpartner jedoch nur geringe Bedeutung zu.
Bereits im ersten Lebenshalbjahr (Unterkapitel 14.6.1) konnten männliche und weibliche Individuen nach ihrem Verhalten eindeutig voneinander unterschieden werden, Männchen spielten mehr, Weibchen waren häufiger Empfänger pflegerischer Verhaltenenweisen.73 Im zweiten Lebenshalbjahr (Unterkapitel 14.6.2) wurden diese Unterschiede noch auffälliger, Weibchen putzten nun auch aktiv signifikant häufiger andere Gruppenmitglieder. Mit diesen Befunden hatten wir belegt, dass für das unterschiedliche Verhalten von Männchen und Weibchen ein Einfluss anderer Gruppenmitglieder nicht notwendig ist. Im zweiten Lebenshalbjahr war zudem erkennbar, dass alle Individuen - wenn auch nur geringfügig - für spielerische Aktivitäten männliche und für pflegerische Aktivitäten weibliche Sozialpartner bevorzugten. Nun wollen wir prüfen, ob mit zunehmendem Alter diese Unterschiede noch deutlicher werden, und berichten über die ersten sechs Lebensjahre der handaufgezogenen Makaken.74 Dabei beschränken wir uns auf wenige, häufig zu beobachtende Verhaltensweisen, nämlich auf Kampfspiel, Kontaktsitzen und Soziale Körperpflege. Darüber hinaus untersuchen wir, ob die jungen Makaken für bestimmte Verhaltensweisen Sozialpartner des einen oder anderen Geschlechtes bevorzugen.
Versuchstiere Im Dezember 1984 ließen wir beide „peer“-Gruppen zusammen. Der Gesamtgruppe stand von diesem Zeitpunkt an eine aus unterteilbaren Innen- und Außenkäfigen bestehende Käfiganlage von insgesamt etwa 30 qm zur Verfügung. Temperatur (27 Grad C) und Luftfeuchtigkeit (60 %) blieben im Innenbereich mehr oder weniger konstant, das Licht wurde von 7.00 bis 19.00 Uhr geschaltet.
Beobachtungsbedingungen Während der Beobachtungen stand den Makaken nur die Innenanlage zur Verfügung, so dass sich dem Blick des zentral vor dem Haltungskäfig sitzenden Beobachters keines der Tiere entziehen konnte. Bei den täglich 30minütigen Beobachtungen protokollierte der jeweilige trainierte Beobachter alle Interaktionen der Sozialgruppe nach vorgegebenem Katalog.
Datenmaterial Bedingt durch die neue Gruppenzusammensetzung, die neue Beobachtungsmethodik (statt Einzeltierbeobachtung nun Gruppenbeobachtung) sind die ab Dezember 1984 erhobenen Daten nicht direkt mit den vorher erhobenen vergleichbar. Auch beruhen die Ergebnisse der einzelnen Lebensjahre auf einer unterschiedlichen Anzahl von Individuen. Eine weitere Ausweitung dieses speziellen Datenmaterials ist nicht möglich, da in dieser Sozialgruppe in der Gruppe geborene Kinder heranwuchsen, so dass sich der Sozialkreis der Individuen änderte (s. u.).
Befundauswertung, Darstellung der Ergebnisse Wir geben an, wie häufig für männliche und weibliche Individuen im Mittel die jeweilige Verhaltensweise während des entsprechenden Lebensjahres protokolliert wurde. Zudem haben wir für das dritte bis sechste Lebensjahr analysiert, inwieweit dem Geschlecht der Sozialpartner Bedeutung zukommt, und geben den mittleren Anteil der männlichen und weiblichen Sozialpartner für Männchen und Weibchen getrennt an. Es sei dabei betont, dass während der gesamten Beobachtungen, auf denen diese Analyse beruht, alle Individuen die Wahl zwischen mehreren Männchen und Weibchen hatten. Alle Daten wurden an der gleichen Gruppe erhoben, die Daten der einzelnen Individuen aber unabhängig voneinander ausgewertet.
Ergebnis
Die bereits in den ersten beiden Lebenshalbjahren auffälligen Unterschiede zwischen
den Geschlechtern, Männchen spielen mehr, Weibchen sind häufiger an pflegerischen
Aktivitäten beteiligt, werden im zweiten Lebensjahr noch auffälliger, sie sind
für die Verhaltensweisen Kampfspiel und soziale Körperpflege durchgängig
statistisch zu sichern. Wie in den ersten beiden Lebenshalbjahren können
wir generell bei dem Kontaktsitzen keine geschlechtstypischen Unterschiede
sichern.
Im zweiten Lebensjahr nimmt die Häufigkeit des Kampfspieles der Männchen
hochsignifikant zu, Weibchen dagegen spielen genauso häufig wie im ersten Lebensjahr.
Der Unterschied zwischen den Geschlechtern in der Häufigkeit des Spielens lässt sich
auch in den folgenden Lebensjahren durchgängig sichern, geht dann aber im 6.
Lebensjahr „verloren“, spielen doch alle Individuen mit zunehmendem Alter immer
seltener.
Individuen beiderlei Geschlechtes putzen im zweiten Lebensjahr andere
Gruppenmitglieder häufiger als im ersten, Weibchen zudem häufiger als Männchen.
Vom dritten Lebensjahr an ist zu erkennen, dass die Häufigkeit der sozialen
Körperpflege durchgängig vom dritten zum sechsten Lebensjahr zunimmt und dass
Weibchen häufiger putzen als Männchen, doch lassen sich diese Unterschiede nicht
mehr statistisch sichern.
Nämliches gilt für die passive soziale Körperpflege, eine Zunahme vom ersten zum
zweiten Lebensjahr mit auffälligen geschlechtstypischen Unterschieden - Weibchen sind
häufiger Empfänger pflegerischer Aktivitäten - und ein Verlust dieser Unterschiede
vom dritteln Lebensjahr an. Es ist dabei bemerkenswert, wenngleich nicht gesichert,
dass Männchen vom 5. Lebensjahr an häufiger geputzt werden als Weibchen. Bei dem
Kontaktsitzen lassen sich Unterschiede zwischen den Geschlechtern nur im 2.
Lebensjahr sichern.
Insofern sind bei den quantitativen Befunden Unterschiede zwischen den
Geschlechtern mit zunehmendem Alter nicht mehr aufzeigbar. Ab dem 3.
Lebensjahr war jedoch auffällig, dass die jungen Javanermakaken bei der Wahl ihrer
Sozialpartner bestimmte Individuen des eigenen oder des anderen Geschlechtes
bevorzugten.
Für das Kampfspiel wählten Männchen und Weibchen bevorzugt männliche Partner.
Dieses Präferensverhalten konnte durchgängig gesichert werden.
Bei der aktiven sozialen Körperpflege ist zu erkennen, dass Männchen Individuen
beiderlei Geschlechtes etwa genauso häufig putzten (mit einer geringen Bevorzugung
gleichgeschlechtlicher Partner), Weibchen dagegen bevorzugten weibliche Partner, was
sich aber nur im sechsten Lebensjahr sichern ließ.
Als passive Putzpartner waren Individuen beiderlei Geschlechtes durchgängig für
weibliche Sozialpartner attraktiver. Dies ließ sich für Männchen im 3., 4. und 6.
Lebensjahr und für Weibchen vom 4. Lebensjahr an gut sichern.
Bei dem Kontaktsitzen dagegen waren gleichgeschlechtliche Partner besonders
attraktiv, besonders auffällig bei den weiblichen Gruppenmitgliedern. Hier
ließ sich die Bevorzugung weiblicher Individuen auch vom 4. Lebensjahr an
sichern.
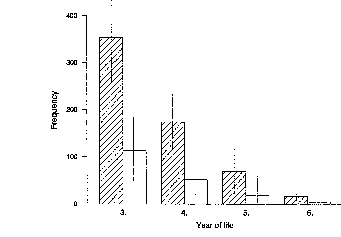
Diskussion Durchgängig werden für nahezu alle Primatenarten auffallende geschlechtstypische Unterschiede im Verhalten berichtet, Weibchen sind häufiger an pflegerischen Aktionen beteiligt. Nach den bisherigen Befunden an den „peer“-aufgezogenen Javanermakaken waren diese geschlechtstypischen Unterschiede bereits im ersten halben Lebensjahr belegbar und nahmen dann mit zunehmender lokomotorischer Reifung erwartungsgemäß zu. Da die jungen Javanermakaken bereits am ersten Lebenstag aus ihrer Geburtsgruppe entfernt und nur mit mehr oder weniger gleichalten Individuen zusammengehalten wurden, ihnen also ältere Gruppenmitglieder nicht als Sozialpartner zur Verfügung standen, konnten wir belegen, dass für diese Unterschiede im Verhalten Lernprozesse nicht Voraussetzung sind. Im Hinblick auf die generellen Befunde an Tierprimaten und den speziellen an unseren Javanermakaken war es insofern unerwartet, dass diese Unterschiede mit zunehmendem Alter wieder (scheinbar) verlorengehen. Berücksichtigen wir nur die quantitativen Befunde, so lassen sich Männchen und Weibchen nach ihrem Verhalten mit zunehmendem Alter nicht mehr unterscheiden. Bei gleichbleibender Häufigkeit positiv-sozialer Interaktionen insgesamt nehmen besonders auffällig die für Männchen charakteristischen spielerischen Interaktionen ab. Der Rückgang in der Häufigkeit des Spielens vom dritten bis zum sechsten Lebensjahr ist gut gesichert.
Der Befund, dass mit zunehmendem Alter die Spielhäufigkeit abnimmt und dass
Adulte überhaupt nicht mehr spielen, gilt aber keineswegs für größere Sozialgruppen
des Javanermakaken mit Tieren aller Altersbereiche. Das Soziale Spiel adulter
Männchen, bzw. auch des Alpha-Männchens, ist vielmehr in allen von uns gehaltenen
Sozialgruppen tatsächlich zu beobachten. Dass in der „peer“- Gruppe von Männchen
und Weibchen als Spielpartner Männchen gewählt wurden, steht dagegen in
guter Übereinstimmung mit zahlreichen Beobachtungen an Sozialgruppen des
Javanermakaken und weiteren von uns gehaltenen Primatenarten. Wahrscheinlich kann
die höhere Attraktivität von Männchen als Partner für das Soziale Spiel als
„Primatenergebnis“ gelten.
Unsere Befunde zum Kontaktsitzen75
zeigen, dass mit zunehmendem Alter der „peers“ offensichtlich gleichgeschlechtliche
Untergruppen gebildet werden, was den zahlreichen Beobachtungen an Makakengruppen
im Labor und im Freiland entspricht. Dennoch widersprechen sie dem bisherigen
Wissensstand. Wie bereits in mehreren Teilen dieser Arbeit betont, ist ein generelles
Ergebnis bei Primaten, dass Weibchen mehr putzen als Männchen und häufiger in
engem Körperkontakt zusammensitzen. Offensichtlich konnten wir letzteres für die
„peer“-aufgezogenen Javanermakaken nicht belegen.
Fragen wir uns nach den Gründen dieser abweichenden Befunde, so war das
Charakteristikum der „peer“ Gruppe das Fehlen jüngerer Gruppenmitglieder. Diese
rufen nach den bisherigen Befunden pflegerische Aktivitäten der Weibchen hervor,
sitzen mit diesen in engem Körperkontakt und dienen ebenfalls als Spielpartner für
adulte Männchen (eigene Beobachtungen). Diese Annahme wird durch Befunde an
derselben „peer“- Gruppe bestätigt. Nach der Geburt von Jungtieren waren diese für
Weibchen Partner bei dem Kontaktsitzen und Ziel pflegerischer Aktivitäten, für
Männchen dagegen Spielpartner. Insofern sind geschlechtstypische Unterschiede
im Verhalten auch abhängig vom sozialen Umfeld. Sie werden bei älteren
Gruppenmitgliedern nur auffällig, wenn entsprechende jüngere Sozialpartner als
Empfänger vorhanden sind. Demnach wird auch verständlich, warum einige Autoren
annehmen, dass das unterschiedliche Verhalten von Männchen und Weibchen von
Erziehungsprozessen abhängig ist, spielen doch tatsächlich nur Männchen mit den
Kindern, Weibchen pflegen sie. Die vielfach beschriebene Orientierung an
gleichgeschlechtlichen Sozialpartnern könnte - so die Annahme - auch das Übernehmen
des Verhaltens des Sozialpartners bedingen. Unsere Befunde zeigen nun eindeutig, dass
diese Annahme falsch ist, für die unterschiedliche Orientierung von Männchen
und Weibchen ist eine Tradierung nicht notwendig. Offensichtlich zeigen dies
auch Primaten, die ohne die Erfahrung einer Sozialgruppe aus Tieren aller
Altersbereiche, insbesondere ohne Mutter, aufgezogen wurden. Die Orientierung zu
Individuen eines bestimmten Geschlechtes lässt zudem geschlechtstypische
Unterschiede nicht reifen, vielmehr ähneln sich Männchen und Weibchen
zunehmend im Verhalten bei der Abwesenheit jüngerer Gruppenmitglieder.
Diese so auffälligen Unterschiede gehen aber keineswegs „verloren“, sie bleiben
vielmehr latent vorhanden und sind stets abrufbare Qualitäten der jeweiligen
Geschlechter.
Neben den hier besprochenen Verhaltensweisen wird in der Literatur auch die
Häufigkeit des agonistischen Verhaltens als Merkmal der Geschlechterunterscheidung
angeführt, so sollen Männchen aggressiver als Weibchen sein, was wir nicht bestätigen
konnten. Tatsächlich richten in jeder Sozialgruppe der hierarchisch organisierten
Makakengesellschaft ranghöhere Individuen agonistisches Verhalten gegen
rangniedrigere. Soweit alle oder fast alle Männchen ranghöher sind als Weibchen, wie
es für einige Species beschrieben wird, ist demnach auch zu erwarten, dass
Männchen häufiger als Weibchen aggressiv zu anderen Gruppenmitgliedern
sind.
Bei unseren Javanermakaken, bei denen nach unseren Beobachtungen zumindest das
Alpha-Weibchen alle anderen Gruppenmitglieder mit Ausnahme des Alpha-Männchens
dominiert, finden wir keine entsprechenden Belege. In der „peer“Gruppe waren sogar
im Mittel die (mehrheitlich auch ranghöheren) Weibchen aggressiver als die
Männchen.
Der Einfluss jüngerer Gruppenmitglieder
Nach Abbruch der Befunderhebungen an den noch kinderlosen
„peers“- in der Sozialgruppe wuchsen nun auch Jungtiere
heran76 -
erhoben wir erneut Daten an dieser Gruppe, als die vier in der Gruppe aufwachsenden
Jungtiere, zwei Männchen und zwei Weibchen, nahezu ein Jahr alt waren
([196]).
Es ist nun statistisch signifikant, dass die handaufgezogenen adulten Weibchen
häufiger das Sitzen in engem Körperkontakt zeigten als Männchen. In Gegegensatz
zu den Befunden der vorhergehenden sechs Jahre, in denen in keinem Jahr
geschlechtspezifische Unterschiede aufzeigbar waren, konnten bei dem Vorhandensein
von Kindern diese Unterschiede auch bei den „peers“ belegt werden.
Betrachten wir das Kampfspiel, dann wird nun auffällig, dass die handaufgezogenen
Männchen wieder signifikant häufiger spielten als die handaufgezogenen Weibchen,
dabei spielten sie nahezu ausschließlich mit den Kindern.
Die Untersuchungen an ohne Kontakt zu Müttern, Geschwistern und älteren Artgenossen in einer Spielgruppe („peer-group“) aufgezogenen Javanermakaken haben für die ersten sechs Lebensjahre erbracht:
1. Männchen spielen mehr als Weibchen. Dieser Unterschied kann ab dem sechsten Lebensjahr nicht mehr gesichert werden, da Männchen und Weibchen in dieser Altersphase kein soziales Spiel mehr zeigen.
2. Die Häufigkeit der Sozialen Körperpflege nimmt bei beiden Geschlechtern mit zunehmendem Alter zu, Weibchen putzen zwar durchgängig häufiger andere Sozialpartner als Männchen, doch ist dieser Unterschied ab dem dritten Lebensjahr nur noch gering.
3. Männchen werden mit zunehmendem Alter als passive Putzpartner attraktiver.
4. Bei der Häufigkeit des Kontaktsitzens können über die gesamte beobachtete Lebensphase keine Unterschiede zwischen den Geschlechtern gesichert werden. Die Befunde zum Kontaktsitzen weisen auf gleichgeschlechtliche Untergruppenbildungen hin.
5. Für das Soziale Spiel wählen alle Makaken unabhängig vom Geschlecht als Sozialpartner vornehmlich Männchen.
6. Unterschiede im Verhalten zwischen den Geschlechtern nehmen anfänglich mit zunehmendem Alter zu, später dann wieder ab. Im sechsten Lebensjahr können Männchen und Weibchen nach der Häufigkeit der hier betrachteten Verhaltensweisen (Kampfspiel, Kontaktsitzen, Soziale Körperpflege) nicht mehr unterschieden werden. Hierfür dürfte das Fehlen jüngerer Gruppenmitglieder verantwortlich sein.
7. Nur bei dem Anwesendsein von Kindern können die Unterschiede zwischen den
Geschlechtern detektiert werden. Männchen spielen mit jüngeren Gruppenmitgliedern,
Weibchen dagegen sitzen mit ihnen in engem Körperkontakt und putzen sie.
Die Kontrollbeobachtungen hielt ich für notwendig, da unsere
Ergebnisse - dem Alter kommt hohe Bedeutung zu, weit höhere als
dem Geschlecht - nach den Literaturdaten eigentlich nicht zu erwarten
waren.77
Wir wollten daher selber entsprechende Experimentalgruppen beobachten, um zu
prüfen, ob unsere Ergebnisse nicht doch generalisiert werden können.
Durch das freundliche Entgegenkommen meines amerikanischen Freundes Stephen J.
Suomi und seinen Mitarbeitern durften wir in Madison, Wisconsin, sieben
„peer-groups“ mit insgesamt 30 Individuen untersuchen, wobei in der Regel
eine Gruppe aus vier Tieren bestand. Zwangsläufig war hierdurch natürlich
eine so hohe Variabilität wie in unseren - größeren - „peer- groups“ nicht zu
erwarten.
Neben Daten zur Änderung des sozialen Beziehungssystems in Abhängigkeit von Alter
und Geschlecht fanden wir, dass auch bei dem Rhesusaffen für die Rangposition des
Individuums dem Alter hohe Bedeutung zukam, ja dass das Alter der Parameter war,
der am besten mit dem Rang korrelierte. Zudem fanden wir auch, dass dem Geschlecht
relativ geringe Bedeutung zukommt, dass also keineswegs Männchen generell
ranghöher sind als Weibchen, wie nach den Literaturbefunden zu erwarten. Wir
erhielten also die gleichen Ergebnisse wie in Kassel, fanden aber hier auch, dass
Abweichungen von dieser altersbedingten Rangordnung zu beobachten sind.
Tabelle 14.8 gibt die Zusammensetzungen der verschiedenen Gruppen nebst der von
uns gefundenen Rangordnung an. Da die Affen in Madison bei Geburt eine
Markierung, entsprechend der Geburtenfolge, erhalten, die aus einer Kombination von
Buchstaben und Ziffern besteht, wobei innerhalb einer Buchstabenkombination ältere
Individuen eine niedrigere Nummer haben als jüngere, können wir erkennen, dass
einige Individuen einen anderen Rangplatz haben, als nach dem Alter zu
erwarten.
Zur Analyse müssen wir aber bedenken, dass es in jeder Makakengruppe im
Freiland und im Labor jeweils ein Alpha-Männchen und ein Alpha-Weibchen
gibt, so dass zwangsläufig die ersten beiden Rangpositionen in einer Gruppe
von diesen eingenommen werden müssen. Berücksichtigen wir also zusätzlich
das Geschlecht der Individuen, wird deutlich, dass in Gruppe 1 AH 48 das
einzige Männchen ist. Auch AH 83 in Gruppe 2 ist das einzige Männchen,
entsprechendes gilt für die Weibchen AJ 81 in Gruppe 5 und AJ 83 in Gruppe 6.
Die abweichenden Rangpositionen dieser vier Tiere können uns also nicht
verwundern.
Auffällig wäre demnach eher, dass weder AH 71 in Gruppe 2 noch AK 63 in Gruppe 7
die weibliche Alpha-Rolle wahrnehmen.
Beachten wir zusätzlich die Geburtstermine der einzelnen „peers“ genauer, dann
finden wir, dass AH 43 und AH 42 von Gruppe 1 am gleichen Tag geboren sind. bzw.
dass der Geburtenabstand zwischen AH 57 und AH 54 aus Gruppe 2 nur zwei Tage
beträgt. Insofern widersprechen die Plätze von AH 43 und AH 57 nicht der Annahme
einer Altersabhängigkeit.
Von den insgesamt 30 Tieren folgen also nur 1/6. nämlich AH 63, AJ 76, AK 62.
AK 64 und AK 63, nicht der Grundannahme einer Altersabhängigkeit der
Rangordnung.
Im Gegensatz zu unseren „peers“ sind leider Daten über das soziale Beziehungsgefüge
der Eltern nicht vorhanden und auch nicht ermittelbar, da diese in der Regel alleine
gehalten werden. Bekannt aber ist für jedes Individuum sowohl der Vater als auch die
Mutter, so dass wir wissen. dass zahlreiche Stiefgeschwister unter den 30 „peers“ sind.
Hierbei handelt es sich jeweils um Stiefgeschwister väterlicherseits. Wir haben daher
auch die Väter für unsere „peers“ angegeben. Betrachten wir die Männchen,
die für mehr als ein Junges verantwortlich sind, so erkennen wir, dass Q 96
nur hochrangige Kinder hat. Das gleiche gilt für S 30 und indirekt auch für
Clancy, dessen beiden Söhne jeweils ihren Rang in der Gruppe verbessert
haben.
Für U 19 hingegen ist auffällig, dass seine Kinder sich offensichtlich schlecht
durchsetzen. Auch 824 hat nur niederrangige Kinder, die Kinder von 1581 haben beide
nicht ihren altersgemäßen Rangplatz in der Gruppe.
Nicht eindeutig sind nur die Befunde für die Väter X 76 und R 11, haben diese doch
sowohl hochrangige als auch niederrangige Kinder. Bedenken wir jedoch, dass das
hochrangige Kind, also AH 81 und AJ 83, jeweils das einzige Weibchen seiner
„peer-group“ ist, müsste man einen hohen Rangplatz eigentlich sogar erwarten.
Insofern widersprechen auch die Daten dieser Männchen nicht der Vermutung, dass
dem Vater Bedeutung zukommen könnte.
Auch der Vater könnte für den Rangplatz seiner Kinder verantwortlich sein. Neben
den Eigenschaften der Mutter, was wir in Kassel belegt haben, kommt demnach wohl
auch noch den unbestimmten Eigenschaften des Vaters bei der Einnahme der späteren
Rangposition Bedeutung zu.
Im Gegensatz zu weiblichen Nachkommen verlassen männliche
Individuen die Geburtsgruppe und versuchen, in eine andere Gruppe
einzuwandern.78
Statt nun die natürliche Situation zu kopieren bzw. statt die jungen Männchen
zu separieren, verzichteten wir auf die Emigration, um Informationen über
die langfristigen Verwandtschaftsbeziehungen der Männchen zu gewinnen.
Wir wollten prüfen, ob ein Männchen in seiner Geburtsgruppe bleiben kann
oder ob der Emigrationspozess obligatorisch ist. Dieser Versuch sollte uns
Antworten über die Grenzen der Familienstrukturen in einer Makakengesellschaft
geben.
Als erstes konnten wir belegen, dass Männchen - unabhängig vom Alter - in der
Geburtsgruppe bleiben können. Sie unterhielten enge Kontakte zu Individuen der
Männchen-Untergruppe, also mit den Individuen, mit denen sie vornehmlich in ihrem
bisherigen Leben interagiert hatten. Sie zeigten submissives Verhalten gegenüber dem
Alpha-Männchen und verhinderten so, Ziel agonistischer Auseinandersetzungen zu
werden. Ende 1983 lebten zehn adulte (bzw. fast adulte) Männchen friedlich in der
Sozialgruppe, nämlich das Alpha-Männchen (Nikita), drei mehr als sieben Jahre alte
und sechs mehr als sechs Jahre alte Männchen. Jedoch änderte sich die Situation
völlig, als eines der jungen Männchen das Alpha-Männchen attackierte und den
Konflikt gewann. Da der Kampf in den frühen Morgenstunden stattfand und nicht
beobachtet wurde, können wir nicht entscheiden, ob dieser Kampf zufällig entstand, ob
das Alpha-Männchen möglicherweise nicht adäquat antwortete oder ob Nikita direkt
angegriffen wurde. Auf jeden Fall verlor er seine Position und musste separiert
werden.79
Das Ergebnis dieses Prozesses war, dass eines der sechs Jahre alten Männchen
(Majo), der Sohn von Mecki, die Alpha-Rolle übernahm und ohne
irgendeine Auseinandersetzung von allen Gruppenmitgliedern akzeptiert
wurde.80
Ein Alpha-Männchen muss eine unangefochtene Postion innerhalb seiner
Sozialgruppe haben. Diese erreicht er, wie ich ausführlich in Unterkapitel
14.2 beschrieben habe, durch das Bedrohen aller anderen Gruppenmitglieder.
Zudem erreicht er es durch das Beißen jedes anderen Gruppenmitgliedes in den
Rücken, bis diese jeglichen Widerstand gegenüber dem Alpha-Männchen aufgeben.
Dieses Verhalten zeigte nun Majo gegenüber allen anderen Gruppenmitgliedern. Fast
unerwartet versuchte keines der Individuen sich dagegen zu wehren.
Das neue Alpha-Männchen wurde aber immer wieder von einem der alten Weibchen
angegriffen, immer in Situationen, in denen Majo eines der anderen adulten Männchen
angriff. Die genaue Analyse der Daten erbrachte, dass es in allen Fällen die Mutter des
angegriffenen Männchens war.
Diese Beobachtungen erlauben die Feststellung, dass Weibchen ihre erwachsenen Söhne
gegenüber dem Alpha-Männchen unterstützen. Dies ist besonders bemerkenswert, da
die Mütter weder ihre Töchter noch ihre jüngeren Söhne in irgendeiner Weise
verteidigten. Insofern können wir erneut fragen, warum heranwachsende Männchen
ihre Geburtsgruppe verlassen müssen. Ohne deren Emigration funktioniert das System
der Makakengesellschaft nicht, da die Söhne-Mütter sich nicht widerstandslos
unterordnen.
Unabhängig von diesen Beobachtungen ging Majo relativ systematisch vor und
unterdrückte sukzessive ein Weibchen nach dem anderen. Drei Monate nach dem
Erreichen der Alpha-Position nahm sich Majo das Weibchen Blonda vor, dabei wurde
diese leicht verletzt. Am Morgen des folgenden Tages war Majo schwer verletzt. Er war
aber offensichtlich siegreich, saß auf einem der höher gelegenen Bretter und zeigte
keinerlei submissives Verhalten gegenüber irgendeinem Gruppenmitglied. Der Rest
der Gruppe wurde dominiert von den drei acht, sieben und fünf Jahre alten
Blonda-Söhnen (Berni, Ernst, Schmidt), die gemeinsam alle anderen Gruppenmitglieder
attackierten.81
Vergleichbare - wenngleich nicht so dramatische - Beobachtungen konnten wir in der
Folgezeit machen. Sobald das Alpha-Männchen die Mutter eines anderen adulten
Männchens angriff, antwortete der Sohn mit verhaltenem Widerstand. So bestieg er z.
B. eines der anderen Weibchen in der Gegenwart des Alpha-Männchens, was einer
Herausforderung gleichkommt.
Unabhängig von diesen Beobachtungen wurde der enge Kontakt zwischen Mutter und
erwachsenem Sohn auch während des normalen Alltags offensichtlich. Auch wenn das
entsprechende Männchen nur leicht verletzt war, suchte es seine Mutter auf, um sich
putzen zu lassen. Zudem muss darauf hingewiesen werden, dass die erwachsenen Söhne
neben den Kontakten zur eigenen Mutter keine engeren Kontakte zu einem der
anderen adulten Weibchen unterhielten. Unabhängig von der Feststellung, dass das
soziale Leben ohne Emigration der Männchen nicht funktioniert, belegten unsere
Beobachtungen, dass Mutter-Kind-Beziehungen stabil bleiben, was wir nun auch für
Söhne belegen konnten.
Ich habe bisher nicht erwähnt, dass ich im Laufe der Jahre zahlreiche weitere Javanermakaken importieren musste, um den Bedarf der Forschungsanliegen von Werner Meinel zu genügen. Die meisten Individuen lebten nur kurz in unserer Haltung. Von diesen zahlreichen Individuen sind nur zwei 1983 importierte Weibchen, Heda und Isolde, von Wichtigkeit. Wir hielten sie - wie auch weitere Makaken - in der neu gebildeten Nikita-Gruppe, deren Interaktionen ebenfalls kontinuierlich protokolliert wurde, wenngleich ich über die Befunde an dieser Sozialgruppe hier nicht berichte.82 In dieser Gruppe belegten beide Individuen, dass sie die Qualitäten haben, die ranghohe Weibchen auszeichnen. Zudem übernahmen wir 1983 die vier Mütter unserer ersten handaufgezogenen Javanermakaken und testeten diese vier umfänglich. Danach war Laurina, die Mutter von Laura, als einziges der vier Weibchen als ranghoch anzusprechen. Insofern konnten wir Laura auch als Tochter eines ranghohen Weibchens bezeichnen, bzw. Heda und Isolde als ranghohe Weibchen für weitere Experimente verwenden.
Die ersten Makaken, die die neue Primatenstation bezogen, waren unsere handaufgezogenen Javanermakaken. Vorher hatten diese - wie bereits berichtet - in meinem Arbeitszimmer im sogenannten Aufbau- und Verfügungszentrum gelebt. Naturgemäß war deren Haltung mit einer gewissen Geruchsbelästigung verbunden. Insofern wurde ich nach dem Auszug von zahlreichen Kolleginnen und Kollegen dankbar angesprochen. Mein warmes feuchtes Arbeitszimmer mit dem reichhaltigen Nahrungsangebot war auch Herberge zahlreicher Schaben, die hier einen optimalen Lebensraum fanden. Dieser war nach dem Auszug der Affen nun plötzlich verloren gegangen. Artgemäß suchten die Schaben nach neuen geeigneten Lebensräumen und wanderten daher aus (letztendlich in den Tod). Ihr Auswandern war freilich mit menschlichem Entsetzen verbunden und mit Auseinandersetzungen zwischen Schabengift-Bekämpfern und -Befürwortern.83
Kollegen, die vorher dankbar das Entfernen der Affen begrüsst hatten,
forderten mich auf, den Raum wieder zu beheizen und dort wieder Affen zu
halten.84
Ich habe bereits berichtet (Unterkapitel 14.1), dass die kleinen Makaken ab dem
zweiten Lebensjahr sukzessive ihre Rangpositionen änderten, „Gewinner“ waren
die Kinder ranghoher Mütter in der Geburtgruppe, „Verlierer“ die Kinder
rangniedriger Weibchen. Besonders spektakulär entwickelte sich der Mecki-Sohn
Martin, das jüngste Jungtier und der einzige Sohn eines ranghohen Weibchens.
Sicherlich auch begünstigt durch sein Alter - als jüngstes Individuum war
Martin besonders attraktiv für die älteren Weibchen seiner Gruppe. In den
der Fusion folgenden Monaten wurde er für uns unerwartet unbestrittenes
Alpha-Männchen und durfte auch als einziges der Männchen alle Weibchen
besteigen. Leider - damals ebenfalls völlig unerwartet - starb Martin bereits
am 21.09.1986. Ich vermute, dass der mit einer Alpha-Position verbundene
Stress (vgl. Unterkapitel 14.11) letztendlich für diesen Verlust verantwortlich
war.

Noch in der letzten Dezemberwoche 1984 zogen wir mit allen Makaken in den Neubau um. Am Tag des Umzuges separierten wir alle nach unserer Definition rangniedrigen Individuen und bildeten so eine neue Gruppe, die nur aus rangniedrigen Individuen bestand. In dieser Gruppe war Max unbestrittenes Alpha-Männchen, ansonsten herrschten chaotische Zustände. 1985 gaben wir dann die zahlreichen Männchen dieser Gruppe ab, so dass Max als einziges Männchen seiner Gruppe verblieb, dennoch herrschte in dieser Gruppe ein „friedliches Chaos“. Max, der das morphologisch kräftigste Männchen unserer Nachzuchtmännchen war, konnte diese Gruppe offensichtlich nicht kontrollieren. Die anderen parallel gehaltenen Sozialgruppen, die Majo-Gruppe mit den übrigen Individuen der ehemaligen Nikita-Gruppe, die neue Nikita-Gruppe und die zusätzlich gebildete Orbi-Gruppe hingegen waren strukturiert. Nach der mühevollen sukzessiven Abgabe fast aller anderen Nachzuchtmännchen bildeten wir am 23.09.1986 zwei weitere neue Gruppen, die neue Majo-Gruppe, die nur aus ranghohen Individuen bestand und die Ernst-Gruppe aus rangmittleren Individuen. Zudem separierten wir (nach den Befunden an der Max-Gruppe, s. u.) die Mütter unserer drei Alpha-Männchen, Mecki (Majo), Blonda (Ernst), Alba (Max), und überführten sie in die Nikita-Gruppe. Ab dem 01.10.1986 wurde dann das Verhalten in diesen drei Gruppen täglich 15 Minuten lang von trainierten Beobachtern protokolliert. Die Befundaufnahme an der Ernst-Gruppe endete leider bereits am 12.11.1987 durch den unerwarteten Tod von Ernst.
Wir bildeten also drei neue Gruppen mit jeweils einem adulten Männchen und sechs
bis acht adulten Weibchen. Mit Ausnahme der beiden Weibchen Heda und Isolde
entstammten alle Individuen der ehemaligen Nikita-Gruppe. Alle Individuen der
Majo-Gruppe waren hochrangige Individuen, die der Ernst-Gruppe mittelrangig und
die der Max-Gruppe niederrangig.
In den ersten sechs Monaten blieben die verwandtschaftlichen Beziehungen erhalten.
Soweit sich über die Mutter verwandte Schwestern in der Gruppe befanden
(Majo-Gruppe: Calva/Cornelia/Angela, Ernst-Gruppe: Flava/Zenia/Tina),
interagierten sie miteinander häufig. Insofern war besonders die geringe
Kontakthäufigkeit zwischen Majo und seinen Schwestern auffällig. Bezüglich der
Mutter-Tochter-Dyaden fiel auf, dass in der Majo- und der Ernst-Gruppe
die Tochter der attraktivste weibliche Sozialpartner der Mutter war (Majo:
Vanda/Frieda, Ernst: Barbara/Olga, und vice-versa. In der Max-Gruppe dagegen
interagierten alle Weibchen häufiger mit nicht (über die Mutter) verwandten
Weibchen als mit ihrer eigenen Mutter/Tochter. Für den einzigen in einem
der Sozialverbände lebenden Sohn, dem Alpha-Männchen Max, blieb die
Mutter der attraktivste soziale Partner. Max interagierte weit seltener mit
adulten (nicht über die Mutter verwandten) Weibchen als die beiden anderen
Alpha-Männchen.
Alle in der Nikita-Gruppe nachgezogenen Weibchen bevorzugten den Kontakt zu
Gleichalten, den Spielpartnern ihrer Kindheit, zudem bevorzugten sie Rangnachbarn
als Sozialpartner. Insofern wurden fast alle zu beobachtenden Präferenzen durch
Verwandtschaft, Alter und Rangposition erwartbar.
Die einzigen sozialen Kontakte, die wir nicht zwingend nach unseren bisherigen
Ergebnissen erwarten konnten, waren diejenigen der zwei neu in die Majo-Gruppe
introduzierten Weibchen (Heda, Isolde), diese unterhielten nämlich enge Kontakte zu
nicht vertrauten Individuen.
Bei der Häufigkeit des Verhaltens Kontaktsitzen fanden wir keine Unterschiede
zwischen den drei Gruppen (im Mittel für alle Gruppenmitglieder: Majo-Gruppe 1347
± 123, Ernst-Gruppe 1262 ± 133, Max-Gruppe 1255 ± 113). Bei dem Zusammensitzen
ohne Körperkontakt zeigten jedoch die Mitglieder der Majo-Gruppe dieses
Verhalten signifikant häufiger (2046 ± 255) als diejenigen der Ernst- (1121 ±
77) und der Max-Gruppe (1158 ± 138). Nämliches galt für das Verhalten
Gehenzu (Majo-Gruppe: 302 ± 35, Ernst-Gruppe: 174 ± 38, Max-Gruppe: 139 ±
22).
Soziale Körperpflege konnte in der Majo-Gruppe (467 ± 86) häufiger beobachtet
werden, was sich gegenüber der Max-Gruppe (139 ± 41) statistisch sichern
ließ.
Vergleichen wir das Verhalten der übrigen Gruppenmitgliedern den drei Alpha-Männchen
gegenüber, so wichen die Gruppenmitglieder häufiger vor Majo (n = 68) als vor
Ernst (n = 43) und häufiger vor Ernst als vor Max (n = 22). Betrachten
wir jedoch andere agonistische Interaktionen - wobei wir zwischen Drohen,
Fliehen, starkem Drohen und Beißen differenziert haben -, zeigte Majo weniger
agonistische Aktivitäten, besonders auffällig bei dem Vergleich von Majo und
Max.
Betrachten wir schließlich die Rangordnung (Tabellen 14.10, 14.13), so erkennen wir
eine lineare Rangordnung in der Majo-Gruppe.
In den Tabellen 14.10 und 14.13 haben wir die Summe aller irgendwie zu beobachtenden agonistischen Interaktionen angegeben (passives Weichen, passives Fliehen, passives Unterwerfungsverhalten (z. B. Zähneblinkeln), aktives Drohen, aktives starkes Drohen und Beißen). In der Majo-Gruppe ist erkennbar, dass eines der Weibchen (Heda) versuchte, ihren Rangplatz zu verbessern, wenngleich erfolglos. Die gleiche Linearität der Rangordnung konnten wir für die Ernst-Gruppe belegen. In der Max-Gruppe hingegen war nach den agonistischen Kontakten Omega ranghöher als Alba. Ein anderes Weibchen (Julia) versuchte das Beta-Weibchen Itta zu dominieren.
Das gleiche Ergebnis erhielten wir bei der Beschränkung auf aktive agonistische
Kontakte (Tabellen 14.11 und 14.14), eine lineare Rangordnung in der Majo- und der
Ernst-Gruppe und eine weit entfernt von einer Linearität existierende Ordnung in der
Max-Gruppe.
Vergleichen wir diese Daten mit denjenigen zur sozialen Körperpflege (Tabellen 14.12
und 14.15), können wir erkennen, dass soziale Körperpflege häufiger in der Majo-
und seltener in der Max-Gruppe zu beobachten war. In den Tabellen 14.12
und 14.15 habe ich die über Verwandtschaft erklärbaren Putzkontakte kursiv
angegeben. Berücksichtigen wir diese und die Erkenntnis, dass Rangnachbarn
engere Kontakte unterhalten, dann ist in der Max-Gruppe nur auffällig, dass
Christa Max, einen Sozialpartner ihrer Kindheit, und Omega Alba häufig
putzte.
In den beiden anderen Gruppen, der Ernst- und der Majo-Gruppe. wurde das
Alpha-Männchen häufig von den beiden höchstrangigen Weibchen geputzt.
Zudem waren diese (das Alpha- und das Beta-Weibchen) attraktive passive
Putzpartner für niederrangige Individuen. Auch die beiden neu in die Majo-Gruppe
introduzierten Weibchen waren als passive Putzpartner auffällig attraktiv. Beide
wurden auch von dem Alpha-Männchen häufig geputzt. Zudem ist sicherlich
bemerkenswert, dass Isolde am häufigsten Heda als passiven Putzpartner
wählte, obwohl sie der Hauptaddressat von Hedas Attacken war (Tabelle 14.14).
Fragen wir nach den Gründen für diese Unterschiede zwischen den Sozialgruppen,
müssen wir als wichtigste Information beachten, dass die Majo-Gruppe nur aus
hochrangigen, die Ernst-Gruppe nur aus mittelrangigen und die Max-Gruppe nur aus
niederrangigen Individuen bestand. Nach unseren Befunden hatten Niederrangige
Defizite bei der Etablierung enger Sozialkontakte. Hochrangige dagegen versuchten
nicht nur Kontakte zu Höherrangigen zu etablieren, sie verfolgten auch eine
Versöhnungsstrategie. Offensichtlich gelten diese Unterschiede auch dann, wenn die
Gruppe nur Individuen des gleichen Rangbereiches enthält. Das bedeutet, dass diese
Unterschiede unabhängig von der aktuellen Position des Individuums in der
Sozialgruppe aufzeigbar sind.
Sicherlich ist es erlaubt anzunehmen, dass die Defizite in der Max-Gruppe auf der
Präsenz seiner Mutter beruhen könnten. Wir hatten daher Alba aus der Gruppe
entnommen (s. o) und parallel zu den Beobachtungen an der Majo- und der
Ernst-Gruppe über weitere sechs Monate Daten an der Max-Gruppe erhoben. Die
Ergebnisse zeigten nahezu identische Befunde, Rangänderungen, Omega und Christa
verbesserten ihren Rangplatz, änderten nichts an dem generellen Befund der
Unfähigkeit, eine lineare Hierarchie zu etablieren.
Auch eine weitere sechsmonatige Befunderhebung an der Majo- und Max-Gruppe belegte erneut die auffälligen Unterschiede zwischen diesen Gruppen (Tabelle 14.17) (Beachte auch die Änderungen in der Rangordnung der Max-Gruppe über die Zeit, Tabellen 14.11,14.16, 14.17) .
Zwar demonstrierten unsere Makaken die Unterschiede zwischen den Gruppen überaus deutlich, doch war mein experimenteller Ansatz - die Voraussetzungen für einen neuerlichen Handaufzuchtversuch zu schaffen - gescheitert, da es nicht gelang zwei gut züchtende Sozialgruppen aufzubauen. 1985 und 1986 züchteten Majo und Max vergleichbar erfolgreich, Majo zeugte 10, Max 7 Jungtiere. Nach den Gruppenneubildungen 1987 bis 1991 wuchsen (bis zum Tod von Max) in der Majo-Gruppe nur 9, in der Max-Gruppe dagegen 17 Jungtiere auf. Nur die Max-Gruppe vermehrte sich hinreichend, in der Majo-Gruppe dagegen züchtete Majo nur mit Heda und Isolde, die übrigen Weibchen waren als Sexualpartnerinnen unattraktiv. Im Nachhinein wäre es sicherlich besser gewesen, ich hätte Majo auch die rangmittleren Weibchen zugesellt, mein Forschungsansatz war zu ehrgeizig gewesen. Die schlechten Zuchterfolge in der Majo-Gruppe sind aus heutiger Sicht gut erklärbar, drei der Weibchen, Calva, Cornelia und Angela, waren Schwestern von Majo und als solche wohl zu vertraut, so dass sie für ihren Bruder als Sexualpartner unattraktiv waren. Zwei weitere Weibchen (Vanda und Frieda) beherrschten perfekt das Verhalten der Konfliktvermeidung und präsentierten ständig, ohne irgendwelche Fluchttendenzen zu zeigen. Dies demotivierte vormals schon Nikita. So blieb auch Frieda das einzige Nachwuchsweibchen unserer Javanermakakenkolonie ohne Nachzuchterfolg. Das Flüchten der Weibchen und das anschließende Verfolgen sind wohl auch bei den Javanermakaken Voraussetzungen, um das Männchen zum Aufsteigen zu motivieren. Dabei fliehen die Weibchen möglicherweise absichtlich, um das Männchen zum Nachfolgen zu bewegen. Ich kann leider hier nur einen anekdotischen Beleg für diese Vermutung anführen. Einmal beobachtete ich, wie ein Weibchen laut schreiend vor dem Alpha-Männchen floh. Dieser verfolgte es aber nicht, vielmehr blieb er ruhig sitzen. Geräuschlos und vorsichtig näherte sich dieses Weibchen dem Alpha-Männchen, um dann sofort wieder laut schreiend zu weichen. Diesen Vorgang konnte ich dann in derselben Beobachtungssitzung mehrfach beobachten.
Noch während der Beobachtungen an der Max-Gruppe starb Vau am 03.01.1987 den
Alterstod. Ihr ganzes Leben lang musste sie sich mit einem niedrigen Rangplatz
zufrieden geben. Am 14.07.1989 verlor Vanda nach mehr als fünfzehn Jahren ihre
Alpha-Position an Heda. Hierfür war wohl die Unterstützung Hedas durch ihre Töchter
Hanne und Herta, möglicherweise auch das Alter von Vanda verantwortlich. Sie starb
am 18.07.1989, bereits einen Tag später verloren wir auch Frieda. Ich vermute,
dass beide psychisch unter dem Rangverlust gelitten haben. Am 04.05.1990
starb Isolde den Alterstod. Schließlich verloren wir Itta am 23.09.1990 durch
Geburtskomplikationen.
Völlig unerwartet starb am 01.03.1992 Max, 6650g schwer. Mit seinem Tod hatten wir
nicht gerechnet, doch ist ein Alter von fünfzehn Jahren ein gutes Alter für ein
Männchen. Wir hatten in unserer Kolonie kein geeignetes (rangniedriges) Männchen
als Ersatz. Unser ältestes neben den beiden Gruppen gehaltenes Männchen war
Theo, ein Sohn des rangmittleren Weibchens Blonda, der zu diesem Zeitpunkt
siebeneinhalb Jahre alt war. In Vorbereitung der neuen Gruppenbildungen
vergesellschafteten wir ihn mit unseren „peer“- Weibchen Sonja und Maria. Am
02.04.1992 gaben wir aus der Max-Gruppe Omega und Jungfrau und aus der
Majo-Gruppe die drei Majo-Schwestern, Calva, Cornelia und Angela, an andere Halter
ab.
Gleichfalls im April 1992 introduzierten wir unsere ranghohen „peer“-Weibchen,
Laura, Carmen und Meike, in die Majo-Gruppe und fusionierten im Mai 1992 die
neu-gebildete Theo-Gruppe mit den restlichen Weibchen der ehemaligen Max-Gruppe.
Beide Gruppenbildungen gelangen. Doch war uns erstmals die Endlichkeit des
Makakenlebens bewusst. Zudem hatten wir in der Majo-Gruppe gelernt, dass zu
große Vertrautheit hinderlich für das spätere Reproduktionsverhalten sein
dürfte. Daher separierten wir am 29.12.1992 Iwan aus der Majo-Gruppe und
Maximilian aus der Theo-Gruppe und hielten beide Männchen gemeinsam in einem
anderen Raum der Primatenstation. Über beide Männchen werde ich noch
berichten.
Auch die neue Majo-Gruppe und die Theo-Gruppe wurden regelmäßig beobachtet,
beispielhaft berichte ich hier wiederum über die Daten von sechs Monaten, die wir ca.
ein Jahr nach Gruppenbildung erhoben haben.
Nicht unerwartet waren in der Majo-Gruppe für Heda ihre Töchter Hanne und Herta attraktivste passive Putzpartner, auch Hanne und Herta putzten ihre Mutter häufiger als eines der anderen Weibchen. In der Theo-Gruppe dagegen putzte zwar Christa ihre Tochter Cora häufiger als andere Weibchen, für Cora waren aber andere Weibchen als passive Putzpartner attraktiver als die eigene Mutter, was an die bereits berichteten Beobachtungen in der Max-Gruppe anschließt. Besonders auffällig im Gruppenvergleich war wiederum der Unterschied in den Beziehungen zum Alpha-Männchen, Majo war als passiver Putzpartner attraktiv und putzte auch selber seine Weibchen, Theo hingegen beteiligte sich an der sozialen Körperpflege sowohl als aktiver als auch als passiver Putzpartner kaum. Bei den aktiven agonistischen Kontakten fällt wiederum auf, dass Theo seine Weibchen häufiger bedrohte als Majo. Zudem ist erneut auffällig, dass die rangniedrigen Weibchen nicht in der Lage sind, eine lineare Rangordnung zu etablieren. Eigentlich unerwartet ist die Rangposition von Heda, ihre von Majo gezeugten Töchter bedrohen auch ihre eigene Mutter und sind höherrangig als diese. Hinweisen möchte ich noch auf die Beziehungen zwischen Alpha-Weibchen (Hanne) und Carmen, Hanne bedrohte Carmen auffällig häufig, „versöhnte“ diese aber wieder. Das erneute „friedliche Chaos“, das Marina Butovskaya85 als „quite tumultuous“ beschrieb ([14]), in der Gruppe der Rangniedrigen begünstigte offensichtlich den Reproduktionserfolg, Theo zeugte 1992 sieben Kinder, Majo nur eines (mit Heda). Auch 1993 zeugte Theo erneut sieben Kinder, in der Majo-Gruppe hingegen wuchsen nur zwei Kinder (von Heda und Hanne) auf, Kinder von Carmen, Laura und Meike starben in den ersten Lebenstagen. 1994 züchtete Theo mit allen neun Weibchen, in der Majo-Gruppe dagegen wurden nur Heda und Hanne schwanger. Zudem mussten wir Carmen separieren. Sie hatte engste Sozialkontakte zu Majo etabliert, griff dann Hanne und Heda massiv an und unterlag. Majo zeugte mit Hanne und Meike Kinder, wobei diesmal das Meike-Kind aufgezogen wurde. Bis zum Tod von Majo am 19.10.1995 wuchsen 1995 in der Theo-Gruppe fünf weitere Jungtiere auf. In der Majo-Gruppe bestätigte sich das unterschiedliche agonistische Verhalten der Hochrangigen, von dem ich bereits in den Unterkapiteln 14.2 und 14.3 berichtet habe, sie kämpften zwar selten, wenn sie sich aber für einen Kampf entschieden haben, kämpften sie richtig und gaben kaum auf.86 Nur aus der Majo-Gruppe mussten wir Individuen nach schweren Verletzungen separieren, in der Theo-Gruppe (wie vorher in der Max-Gruppe) herrschte stets Chaos, doch traten erhebliche Verletzungen über die Jahre nicht auf. Marina Butovskaya beschrieb auch das unterschiedliche Verhalten der beiden Alpha-Männchen87 und der weiblichen Gruppenmitglieder88, sie kam auch zu dem Schluss, dass für diese Unterschiede genetische Faktoren89 verantwortlich sein könnten. Dies hatten wir leider nicht belegen können, stand doch das neuerliche Handaufzuchtexperiment noch aus. Während meiner Zeit in der Primatenstation konnte ich dieses Experiment nur noch vorbereiten, aber nicht mehr durchführen.
Ich habe im Unterkapitel 14.9.5 bereits berichtet, dass wir am 29.12.1992 Iwan und
Maximilian aus ihren Geburtsgruppen separiert hatten. Diese hielten wir
komplikationslos gemeinsam in einer Haltungseinheit. Am 04.08.1994 überführten wir
diese in eine andere Einheit, separierten sie am 11.11.1994 und verpaarten Iwan mit
seiner Halbschwester Herta. Maximilian vergesellschafteten wir mit Amanda,
einer mutteraufgezogenen Tochter von Theo und Elke, und einem weiteren
handaufgezogenen weiblichen Jungtier.
Am 21.04.1995 wurde Vincent geboren, Iwan hatte seine
Zeugungsfähigkeit unter Beweis gestellt. Einen Tag nach dem Ableben von
Majo90 -
am 19.10.1995 - introduzierten wir Iwan, Herta und Vincent in die ehemalige
Majo-Gruppe. Iwan übernahm sofort unangefochten die Rolle des Alpha-Männchens
und kontrollierte problemlos die Gruppe. Wir mussten zwar noch im Oktober 1995
Laura und Meike separieren, doch war nun unsere Gruppe der ranghohen
Individuen auch reproduktiv aktiv, Iwan züchtete mit Hertha, Hanne und
Charlotte sowohl 1996 als auch 1997 erfolgreich unter den Jungtieren waren zwei
Töchter. Insofern konnten wir mittelfristig eine genügende Zahl von Jungtieren
erwarten. 1997 wurde diese interessante Gruppe aus der Primatenstation
entfernt.
Am 12.06.1996 gaben wir das handaufgezogene Weibchen aus der
Maximilian/Amanda-Gruppe ab und überführten Maximilian und Amanda in ein
größeres Gehege der sich nun sukzessive leerenden Primatenstation. Am 24.07.1997
gebar Amanda ihr erstes Kind, auch der Max-Sohn Maximilian, der zu einem großen
Männchen herangewachsen war, hatte seine Zeugungsfähigkeit belegt. Auffällig war in
dieser Gruppe, dass Amanda die Nähe zu Maximilian mied, sie bevorzugte möglichst
weit von ihm entfernt liegende Ruheplätze.
Theo war in seinen letzen Lebensjahren zuckerkrank. Ich diagnostizierte dieses Leiden
nach dem Verhalten der reichlich in seiner Gruppe vorhandenen Jungtiere.
Sobald er urinierte, sprangen diese herbei, um den Urin aufzulecken. Auffällige
Harnaufnahme hatten wir ansonsten bei unseren Makaken nicht beobachtet. Da Theo
trainiert war, den Absperrgang aufzusuchen, konnten wir ihn auch mit Insulin
versorgen. Ich selber war in dieser Zeit - wegen meiner fristlosen Entlassung - nur
partiell anwesend. Hier versorgte ihn mein Doktorand Michael Dulitz täglich
aufopfernd.
In den letzten Jahren der Javanermakakenhaltung wurden diese von meiner
Doktorandin Christiane Möller begleitend beobachtet. Leider konnte Christiane Möller
ihre Doktorarbeit durch die Geschehnisse in der Primatenstation nicht mehr
abschließen. Mir liegen auch keine Unterlagen vor. Möglicherweise werde ich bei einer
Neuauflage dieses Buches - sollte Frau Möller ihre Aufzeichnungen aufbewahrt haben -
hier ausführlicher berichten können. Frau Möller beobachtete im Rahmen ihrer
Diplomarbeit, die mir leider auch nicht (mehr) vorliegt, 1996 auch bereits die
Theo-Gruppe. In dieser lebten u. a. drei 1993 geborene Männchen und zwei 1994
geborene Weibchen (Naomi und Guillian). Während ihrer Beobachtungen stellten
Naomi und Guillian nach meiner Erinnerung das soziale Spiel (Nachlaufen) völlig ein.
Dann gaben wir am 12.06.1996 die drei Männchen planmäßig an andere Halter
ab. Ab diesem Zeitpunkt nahmen die beiden jungen Makakenweibchen das
Nachlaufspiel wieder auf. Diese Beobachtung erlaubt die Hypothese, dass das
rauhe Kampfspiel der Männchen die jungen Weibchen veranlasst, das Spielen
einzustellen.
Theo ging es 1997 zusehends schlechter, auch verlor er zumindest einen Teil seiner
Sehkraft. Da zwei Haltungseinheiten im selben Raum leerstanden, die Iwan-Gruppe
war abgegeben, ebenso die im mittleren Gehege ehemals lebende Totenkopfaffengruppe
(Viktor-Gruppe), wurde ich regelmäßig von Beauftragten der Universität dringenst
aufgefordert, der Theo-Gruppe mehr Platz zu geben. Mein Einwand, man
täte einem fast blinden Männchen, der sich in seinem vertrauten Innen- und
Außengehege optimal orientieren könne, mit mehr Platz keinen Gefallen,
vielmehr würde man diesem schaden, wurde als Einwand eines uneinsichtigen
Halters abgetan, doch konnte ich mich - vorübergehend wieder im Amt -
durchsetzen.
Etwa im August 199791
starb dann Theo. Ich beschloss, Maximilian vor einer Fusion mit den Weibchen der
Theo-Gruppe zu verstärken und integrierte in seine Gruppe auch Sophie und
Christa, die beiden ältestesten Weibchen der ehemaligen Theo-Gruppe. Bei
diesem Experiment verloren wir Sophie (am 30.08.1997). Wie und warum
diese zu Tode kam, kann ich aus der Erinnerung nicht berichten. Maximilian
jedenfalls hatte mit Christa ein Weibchen gefunden, mit dem er gerne und ständig
zusammensaß.
Wohl im Dezember 1997 fusionierten wir dann die Maximilian-Gruppe (Maximilian,
Christa, Amanda und Amanda-N) mit dem Rest der Theo-Gruppe. Es herrschte das
reine Chaos. Maximilian hatte panische Angst vor den neuen Weibchen und floh vor
ihnen. Die Theo-Weibchen dagegen hatten ebenfalls panische Angst vor dem großen
Mann92
und flohen vor diesem. Amanda sah die Chance, den Abstand zu Maximilian
zu vergrößern, und wich in die gleiche Richtung wie die übrigen Weibchen,
wurde aber sofort von den Theo-Weibchen angegriffen und gebissen. Nur
Christa blieb ruhig auf einem oberen Brett sitzen und war für Maximilian ein
verlässlicher Sozialpartner. Bei der Separation von Christa hatten wir sie auch
von ihrem am 01.06.1996 geborenen Sohn Alonso getrennt. Nach der Fusion
suchte dieser seine Mutter sofort auf und wurde von dieser am Bauch getragen.
Als erstes merkte Maximilian, dass er nicht vor den Weibchen fliehen muss, er konnte
unbesorgt und unbedrängt bei Christa und Alonso bleiben. Auch Amanda erkannte,
dass es sinnvoll ist, in der Nähe von Maximilian zu bleiben, entging sie doch so
den Attacken der Weibchen. Es trat eine relativ friedliche Atmosphäre ein,
nur Sonja und Maria konnten offensichtlich nicht begreifen, dass sie ihren
hohen Rangplatz an Christa verloren hatten. Gemeinsam mit Guillian, der
verwaisten Tochter von Sophie, und Heiko saßen sie eng gedrängt mit ihren
Kindern auf dem Boden und drohten Christa (und damit auch Maximilian). Am
11.12.1997 beobachtete ich mit Christiane Möller die Gruppe, wir waren uns einig,
die Fusion habe geklappt. Beim Verlassen des Raumes stürmte Maximilian
auf den Boden und tötete Heiko, den Sohn von Sonja, mit einem Kopfbiß.
Dies war der erste Fall von Infantizid in unserer Makakenkolonie. Nach dem
Tötungsbiß schüttelte sich Maximilian und spuckte, als hätte er irgendetwas Ekliges
gegessen. Am 12.12.1997 fanden wir Maria-N und am 13.12.1997 Guillian durch
Kopfbiss getötet vor. Es spricht alles dafür, dass auch hier Maximilian der Töter
gewesen ist. Er hat Kinder der Weibchen getötet, die sich nicht untergeordnet
haben, zudem das einzige Jungtier, das engste Kontakte zu diesen Weibchen
hielt.
Hätte Maximilian die Primatenliteratur und die vielen klugen Artikel zum Infantizid
bei Primaten gelesen, hätte er statt dem dreijährigen Weibchen Guillian besser
Alonso (und weitere Kinder) getötet, so wuchs Alonso als Stiefsohn unverletzt
heran.
Letzendlich war auch diese Fusion geglückt, wir hatten wiederum eine Gruppe aus nur
nach der Geburt rangniedrigen Individuen. Ich persönlich bin überzeugt, hätten wir
unser Handaufzuchtprojekt nun wiederholt, wären alle Iwan-Kinder im oberen Bereich
der Hierarchie und alle Maximilian-Kinder im unteren Bereich gelandet. Nach meiner
Einschätzung werden die sozialen Eigenschaften, die Persönlichkeiten, die letztendlich
für einen hohen oder einen niedrigen Rang verantwortlich sind, eindeutig vererbt.
Für die unterschiedlichen sozialen Strategien und für die unterschiedlichen
Persönlichkeiten der Individuen habe ich in diesem Unterkapitel zahlreiche Befunde
berichtet.
Bereits mehrfach in diesem Buch habe ich meine Zweifel geäußert, ob Primaten - wie
theoretisch für alle Lebewesen gefordert - Verwandtschaft - gesteuert von ihren Genen -
detektieren könnenen. In einem Review zur Sozialstruktur der Primates ([247]) kam
ich nach gründlicher Diskussion vermeintlicher Belege für das Verwandtenerkennen zu
dem Schluss: „Zusammengefasst dürfte die hohe Bedeutung der verwandtschaftlichen
Beziehungen vor allem auf der Leistung des Individuums beruhen, andere Tiere
individuell anzusprechen, auf das Vermögen, nach dem Grad der Vertrautheit
bestimmte Tiere vor anderen zu bevorzugen, und auf dem dadurch entstehenden
Bewusstsein für die eigene Familie, eigene Untergruppe bzw. eigene Gruppe. ... Dabei
ist es dann unerheblich, ob die sich gegenseitig unterstützenden Tiere wirklich
miteinander verwandt sind, wird durch dieses Verhalten doch das Entstehen
komplexer Sozialgefüge im Labor und im Freiland erst ermöglicht.“ ([247], Seiten
146-147).
Für das Vorhandensein eines ohne individuelles Kennen vorhandenes Erkennen von
Verwandtschaft gab es damals einen wichtigen Beleg. In der Zeitschrift Nature war
1980 eine Arbeit von Wu et al. ([305]) erschienenen mit dem Titel: „Kin preference in
infant Macaca nemestrina“. Diese Arbeit schien damals der Beleg zu sein, dass auch
Primaten Verwandtschaft detektieren können. Der Arbeit kam besonderes Gewicht zu,
hatten die Autoren doch den aufwendigen Review-Prozess der angesehensten
naturwissenschaftlichen Zeitschrift bestanden bzw. kamen die Daten aus einer
renommierten Forschungseinrichtung.
Weiß nun ein Individuum - gesteuert von seinen Genen -, ob es mit einem anderen
verwandt ist oder nicht. Zur Klärung dieser Frage führte Wu gemeinsam mit anderen
Mitarbeitern der Arbeitsgruppe um Sackett - mein Freund Gene P. Sackett war auch
Co-Autor dieser Studie - an sechzehn Schweinsaffen, die kurz nach der Geburt
separiert wurden, Wahlexperimente durch. Sie sperrten die Kinder in den von Sackett
entwickelten Wahlapparat. Von einem Ausgangskäfig aus durften die jungen
Schweinsaffen in drei Abteile blicken. Jeweils durch eine zusätzliche Scheibe getrennt
sass in einem ein väterlichseits verwandtes Halbgeschwister und in dem zweiten ein
nichtverwandtes Individuum gleichen Alters und Geschlechtes wie das Geschwister.
Das dritte Abteil schließlich blieb leer. Nach einer fünfminütigen Orientierungsphase
durften die Jungtiere zehn Minuten lang die gleichzeitig geöffneten Abteile wahlweise
betreten, wobei die jeweilige Zeit des Aufenthaltes genauestens protokolliert
wurde.
Es zeigte sich nun, dass Halbgeschwister vor Nichtverwandten präferiert wurden.
Eindeutig zeigten 13 der 16 Kinder eine solche Präferenz, was sicherlich hochsignifikant
ist. Unabhängig von meinen grundsätzlichen Zweifeln, ob 10 Minuten Wahlversuch
hinreichend sind, um Verwandtenerkennen wirklich zu belegen, habe ich die Daten
genauer betrachtet. Tatsächlich hatten die jungen Schweinsaffen vier Optionen, sie
konnten den Verwandten oder den Nichtverwandten aufsuchen, das leere Abteil oder
aber im Ausgangsort verbleiben. Nun zeigte sich, dass nur sechs der Jungtiere den
Verwandten präferierten, drei präferierten (s. o.) den Nichtverwandten, zwei das leere
Abteil und fünf schließlich den Ausgangsort. Betrachten wir zudem die sechs Belege,
dann fällt bei zwei weiteren auf, dass der Unterschied zwischen Verwandten
und Nichtverwandten nur sehr gering ist. Die Daten von Wu et a. geben also
überzeugend nur an, dass von den sechzehn Kindern vier Halbgeschwister und drei
Nichtverwandte präferierten. Die Resultate konnten mich nicht überzeugen.
Durch unsere beiden „peer“-Gruppen hatten wir das optimale „Ausgangsmaterial“ für
ein klärendes Experiment (vgl. auch [249], [284], [290]). Dieses Experiment war die
Fusion unserer beiden „peer“-Gruppen und die Beobachtungen zum Sozialverhalten
der 20 Makaken.
Es wurde im Dezember 1984 durchgeführt, die Makakenkinder waren zu diesem
Zeitpunkt zwischen 13 und 42 Monaten alt. Die älteren „peers“ wurden sukzessive in
halbstündigem Abstand, beginnend mit dem jüngsten Individuum (Klara)
entgegengesetzt zum Alter in die Gruppe der zwölf jüngeren Individuen gelassen. Die
Beobachtungen begannen am darauffolgenden Tag. Über zehn Tage protokollierten zwei
trainierte Beobachter täglich zwei Stunden lang die Interaktionen der zwanzig
Jungtiere, nämlich alle affiliativen Ereignisse (Kontaktsitzen, soziale Körperpflege,
Zusammensitzen (mit und ohne Körperkontakt), Gehenzu) und alle agonistischen
Episoden.
Die Fusion der beiden Gruppen ermöglichte es jedem Individuum, dyadische
Beziehungen zu (1) Vertrauten-Nichtverwandten, zu (2) Vertrauten-Verwandten, zu (3)
Nichtvertrauten-Nichtverwandten oder zu (4) Nichtvertrauten-Verwandten aufzunehmen.
Vergleichen wir die Ergebnisse zur Vertrautheit und zur Verwandtschaft (wobei wir
hier nicht zwischen den Graden der Verwandtschaft (Halbgeschwister/Vollgeschwister)
differenziert haben), dann wurden 77,8 % der affiliativen sozialen Kontakte -
insgesamt wurden 59579 Kontakte protokolliert - zwischen Vertrauten beobachtet,
d. h. die Javanermakaken präferierten ihre ehemaligen Sozialpartner in den
beiden Ausgangsgruppen. Soweit sie zwischen Vertrauten-Verwandten und
Vertrauten-Nichtverwandten wählen konnten, ergaben sich keine Unterschiede.
Nämliches galt für das Wahlverhalten zu nichtvertrauten Individuen. Sie unterschieden
nicht zwischen nichtvertrauten-verwandten und nichtvertrauten-nichtverwandten
Individuen.
Da dreizehn Makaken auch die Möglichkeit hatten, zu Vollgeschwistern den Kontakt
aufzunehmen, analysierten wir auch diese Variable. Eindeutig interagierte kein
Individuum häufiger mit einem Vollgeschwister als zu erwarten. Offensichtlich beruht
das in Sozialverbänden zu beobachtende Präferenzverhalten gegenüber Verwandten auf
Vertrautheit.
Unsere Ergebnisse entsprachen nicht nur unseren Erwartungen, vielmehr hatte ich
nach meiner Einschätzung ein wichtiges Ergebnis für eine Nature-Publikation erreicht.
Im Nature-Format schrieb ich ein Manuskrpt „Failure of kin-recognition in Macaca
fascicularis“ und reichte diese bei Nature ein. Zu meiner Überraschung wurde diese
abgelehnt, sie wäre nicht von allgemeinen Interesse. Ich habe meine Ergebnisse
dennoch publiziert ([290]), war aber irritiert.
Bei einer Vortragsreise in den USA besuchte ich auch auf Einladung von
Gene P. Sackett das Washington RPRC in Seattle und berichtete in
meinem Vortrag von dieser Antwort. Die Kollegen lachten nur verlegen,
sie hätten selber - begeistert von ihren Ergebnissen - eine umfangreiche
Untersuchung zum Thema durchgeführt und ihre eigenen Daten eindeutig
widerlegt.93
Auch diese Arbeit wurde von Nature abgelehnt, dennoch aber publiziert ([58]).
Insofern drängt sich der Verdacht auf, dass das Ergebnis, eine eindeutige Falsifikation
einer der Grundannahmen der Soziobiologie, nur von geringem Interesse ist. Die
Theorie möglicherweise stützenden Ergebnisse hingegen sind von allgemeinem
Interesse, auch wenn sie eigentlich kein echter Beleg sind. Eine Möglichkeit, diese zu
widerlegen, wird von Nature nicht gegeben. Freilich hat eine Publikation in
einer anderen Zeitschrift weit weniger Gewicht, d. h. der Fehlbefund bleibt als
gesichert erhalten. Seit diesem Zeitpunkt lese ich Nature-Publikationen nur mit
Vorbehalt.
In Unterkapitel 14.7 habe ich meine Kontrollbeobachtungen an Rhesusaffen
(Macaca mulatta) berichtet, die ich durch die freundliche Genehmigung von
Stephen J. Suomi am Harlow-Laboratory in Madison, Wisconsin, durchführen
durfte.
Möglicherweise angeregt auch durch unser kontinuierliches Nachfragen nach
der Vaterschaft der jungen Makaken oder auch unabhängig hiervon führte
ein Mitarbeiter von Stephen J. Suomi, James M. Scanlan, vergleichbare
Untersuchungen an 73 Rhesusaffenjungtieren durch und erzielte vergleichbare
Ergebnisse.94
Er analysierte auch sorgfältig weitere Variablen und und kam - wie wir in Kassel - zu dem
Schluss, dass genetische Faktoren für die soziale Stellung des Individuums verantwortlich sein
dürften.95.
Darüber hinaus erhob James M. Scanlan Daten zur Hormonsekretion an diesen
Rhesusaffen. - Stephen J. Suomi hatte bereits früher auf einer Tagung in
Italien, bei der ich ihn auch ansprach und gleichzeitig die Voraussetzungen für
die Beobachtungen in Wisconsin erkundete, individuelle Unterschiede in der
Hormonsekretion aufgezeigt. Er nannte die eine Gruppe von Individuen nach
meiner Erinnerung „more reactive“, die andere „less reactive“ (vgl. auch
[210]).
Nach Scanlans Befunden konnte man die Kinder der hochrangigen
und die der niederrangigen Männchen eindeutig nach der
ACTH-Sekretion96
diskriminieren.97
Diese Befunde bewogen mich zu untersuchen, ob wir ähnliche Hormon-Befunde bei
unseren Javanermakaken aufzeigen könnten. Für diese - im Vergleich zu unseren
Verhaltensbeobachtungen - teure Untersuchung warb ich Mittel bei der DFG ein, so
dass wir diese 1990 durchführen konnten.
Das Vorgehen war nach den damaligen Literaturangaben vorgegeben, die Makaken
sollten einem definierten Stress ausgesetzt - fünfzehn-minütiger Aufenthalt in einem
Zwangskäfig- und anschließend vor der Blutentnahme betäubt werden. Wir hatten
damals keine Erfahrung mit der sachgerechten Betäubung von Javanermaken, auch
fehlten uns damals die Techniken der Blutentnahme. Durch die großzügige
Unterstützung von Ernst Weinmann, der dem routiniertesten seiner Mitarbeiter in
der Primatenhaltung, Herrn Kraft, gestattete, die Blutentnahmen in Kassel
durchzuführen, konnten wir das Experiment angehen. Herr Kraft war auch geübt in
der Dosierung der Betäubung von Javanermakaken, so dass diese kein Problem
darstellte. Ernst Weinmann stellte mir auch ein geignetes Netz zum Einfangen der
Tiere zur Verfügung, wobei ich das nicht ungefährliche Einfangen und Umsetzen in
unsere hausgebauten Zwangskäfige übernahm. Ein in Vorbereitung des Vorhabens mit
Endokrinologen durchgeführtes Consilium „segnete“ das Vorhaben ab, ein Einfluss
der Betäubung auf die Hormonsekretion konnte nach den Literaturbefunden
ausgeschlossen werden.
Ein kritisches Moment bei der Betäubung von Tieren ist immer die Aufwachphase. In
den relativ kleinen Haltungskäfigen der Behringwerke bzw. des Harlow Primate Lab
oder anderer Primatenzentren konnte das Problem vernachlässigt werden, die
betäubten Individuen wachen in ihren normalen Haltungskäfigen auf. Wir
mussten uns hier wegen unserer relativ hohen Haltungseinheiten eine Alternative
überlegen. Ich bereitete für jedes Individuum eine Bananenkiste vor, die wir nach
der Betäubung in die Haltungsgehege stellten, davon ausgehend, dass die
Javanermakaken erst nach dem vollständgen Aufwachen, sich aus diesen Kisten
befreien könnten.
Das Einfangen klappte problemlos, wobei ich auf das Einfangen von Majo und Max
verzichtet habe, ich befürchtete, beide wären in unseren Zwangskäfigen nicht zu
halten. Die Betäubung und die Blutentnahme klappten dank des Einsatzes von Herrn
Kraft, auch das Verbringen der betäubten Individuen in die Haltungskäfige
wurde schnell und sorgfältig dank meines Mitarbeiter-Teams durchgeführt. Das
Aufbereiten der Blutproben für die Analyse übernahm Michael Herzog, ein
Marburger Medizindoktorand. Nach dem stundenlangen Einsatz atmeten
alle Beteiligten auf, Herr Kraft war von unserer Arbeitsatmoshäre und dem
Einsatzwillen aller Mitarbeiter angetan und bot an, jederzeit wieder nach
Kassel zu kommen, er würde sich auch dafür Urlaub nehmen. Doch ging meine
Planung mit den Bananenkisten nicht auf. Gerade die großen Männchen, die wir
damals noch hielten, befreiten sich unerwartet schnell aus den Kisten und
suchten höher gelegene Ruhebretter auf. Dort torkelten sie umher, so dass ein
Runterfallen vorstellbar war und nicht ausgeschlossen werden konnte. Im
Nachhinein hatten wir in Kassel nicht die geeigneten Voraussetzungen für
diese Untersuchung, hier wären viel kleinere Haltungskäfige Voraussetzung
gewesen. Wir haben zwar kein einziges Individuum durch dieses Experiment
verloren, doch schien mir eine Wiederholung der Prozedur nicht vertretbar zu
sein.
Gewissermaßen „entschädigt“ wurden wir durch die Ergebnisse dieser
Untersuchung ([288]). Unsere hochrangigen Individuen hatten eine
signifikant niedrigere ACTH-Sekretion. Wir hatten Anschluss an die
Untersuchungen der Arbeitsgruppe von Stephen J. Suomi gefunden. Neben der
ACTH-Sekretion untersuchten wir auch diejenige von Cortisol, Adrenalin und
Noradrenalin.98
Nach der Cortisolsekretion konnten wir ebenfalls Ranghohe und Rangniedrige
unterscheiden, die ranghohen Individuen hatten eine signifikant höhere Cortisolsekretion,
nämliches galt für die Adrenalin- und Noradrenalinsekretion, doch ließen sich letztere
Unterschiede nicht sichern.
Eine Wiederholung des Experimentes war - wie schon gesagt - unter unseren
Haltungsbedingungen nicht ratsam. Ich hatte aber während eines USA-Aufenthaltes
auch die Freianlagen des Yerkes RPRC, Atlanta, Georgia, besucht, wo u. a.
Rhesusaffen in fußballfeldgroßen Gehegen gehalten wurden. Hier demonstrierte mir
Irwin S. Bernstein seine Form der Blutentnahme.
Er ging in die Innenanlage, öffnete ein Fenster zum Außengehe und rief „Come in“.
Wie von Geisterhand gesteuert rannten alle Rhesusaffen in die Innenanlage, sprangen
dann sukzessive in einen Gitterkäfig, streckten ein Bein zur Blutentnahme heraus und
suchten dann wieder das Außengelände auf. Meine Mitarbeiterin Cornelia Schäfer-Witt
und ich beschlossen daher, den Versuch zu machen, auch unsere Javanermakaken zu
trainieren.99
Ich ließ ([251]) in den Zentralen Werkstätten der Universität einen fahrbaren Tunnel
bauen, der in jedem der Haltungsgehege installiert werden konnte. Die Haltungsgehege
konnten, wie schon in Kapitel 12 beschrieben, in zwei Abteilungen geteilt werden.
Wir sperrten die Gruppenmitglieder in Abteilung A und gaben ihnen die
Möglichkeit, das Innengehege durch den offenen Tunnel in das Außengehege zu
verlassen.100
Innerhalb von Minuten verließen alle Gruppenmitglieder das Innengehege über diesen
Weg. Am folgenden Tag wiederholten wir dieses Experiment. Nun verließen alle Affen
das Innengehege innerhalb weniger Sekunden. Am dritten Trainingstag verschlossen
wir den Tunnel nach außen (der horizontale Schieber wurde geschlossen), nun
betrat keiner der Makaken diesen. In den folgenden Stunden musste ich jedes
Individuum, auch Majo und Max, einzeln mit dem Netz einfangen und durch
den Verbindungschieber in den Tunnel frei lassen. Hier verschloss Cornelia
Schäfer-Witt den Tunnel und gab anschließend den Weg nach außen frei. Einen
weiteren Tag später suchten fast alle Affen den verschlossenen Tunnel auf. Nur
wenige mussten erneut eingefangen werden. Am fünften Trainingstag war das
Tunnelaufsuchen Routine. Am sechsten Tag lernten sie, im Tunnel separiert zu
werden, am siebenten, dass Blutentnahme unproblematisch ist, sie mussten nur
ein Bein durch das Gitter stecken. Naturgemäß dauerte es etwas Zeit, bis
allen Tieren Blut abgenommen werde konnte. Einige wehrten sich. Da aber
das mittlere Abteil als Zwangskäfig konstruiert war, konnten wir ein Bein
greifen, ohne Gefahr für die menschlichen Betreuer. Wir erreichten also sehr
schnell unser Ziel, Blutentnahmen ohne Betäubung durchführen zu können.
Dabei gewannen wir sogar den Eindruck, dass unsere Makaken das Handling
mochten, besonders auffällig war dies bei den „peer“ aufgezogenen Individuen, sie
schienen den Kontakt zu den Menschen zu genießen. Die Blutentnahme selber
belastete unsere Makaken nicht, was auch verständlich ist, ist doch der ihnen
zugefügte milde Schmerz nicht zu vergleichen mit dem täglichen Stress bei dem
Umgang mit Artgenossen. Probleme entstanden nur bei Änderungen der Routine.
Unsere Prozedur, die ich in Unterkapitel 14.12 ausführlicher beschreibe, war
nämlich die folgende: Wir installierten den Gang nur im mittleren Gehege.
Insofern mussten die Makaken mehrere Innen- und Außengehehe durchqueren,
bevor sie in ihr Innengehege zurück gelangten. Gaben wir ihnen nach dem
Entfernen des Tunnels die Gelegenheit, direkt in ihr Gehege zurückzukehren (ohne
Umweg über das Außengehege), wirkten sie verzweifelt und gestresst. Wir
lernten, dass es geboten ist, auf jegliche unnötige Änderung der Routine zu
verzichten.
Die Blutentnahmen für die Analyse der Hormondaten an den nicht betäubten Affen
übernahm Michael Herzog, bei der Aufarbeitung der Proben für die Analyse wurde er
unterstützt durch Marburger Co-Doktoranden. Cornelia Schäfer-Witt und ich
„stellten“ sozusagen nur das Bein der jeweiligen Individuen zur „Verfügung“,
übernahmen also alle Trainingsaufgaben.
Die Analyse der Daten überraschte uns, erneut konnten wir zwar die ranghohen
und die rangniedrigen Individuen eindeutig nach der Hormonsekretion der
Hypophysen-Nenennieren-Achse unterscheiden, doch erhielten wir entgegengesetzte
Resultate, die ACTH-Werte der Ranghohen waren signifikant höher als bei den
Rangniedrigen, bei der Cortisol-Sekretion hingegen lagen die Cortisol-Werte der
Rangniedrigen signifikant über denen der Ranghohen. Erneut ergab die Analyse der
Adrenalin-und Noradrenalin-Sekretion keinen signifikanten Unterschied zwischen
beiden Sozialgruppen. In beiden Experimenten (betäubte und nichtbetäubte
Individuen) waren die Mittelwerte der Ranghohen bei Adrenalin und Noradrenalin
höher als bei den Rangiedrigen, die hohe Streuung der Daten erlaubte aber keine
Sicherung dieses Befundes.
Wir haben daher die Individualwerte genauer betrachtet und konnten erkennen, dass
bei beiden Experimenten die jeweiligen Alpha-Weibchen signifikant höhere Werte
hatten als die übrigen Weibchen ihrer jeweiligen Gruppe. Das gleiche galt für Majo
und Max, bei denen uns nur die Werte als unbetäubte Individuen zur Verfügung
standen. Die hohen Werte der Alpha-Tiere waren für die Streuung der Werte
verantwortlich.
Nach unseren vorläufigen Befunden ist die Hypothese erlaubt, dass sich der
„Geburtsrang“, also die unterschiedlichen Persönlichkeiten der Hochrangigen und der
Niederrangigen an der Sekretion von Cortisol und ACTHstichACTH festmachen lässt,
der aktuelle Rang - unabhängig von den unterschiedlichen Persönlichkeiten -
manifestiert sich in der Adrenalin- und Noradrenalin-Sekretion.
Die Unterschiede zwischen beiden Experimenten scheinen uns leicht erklärbar zu sein.
Die niederrangigen Individuen haben per se eine höhere Cortisol-Sekretion als die
hochrangigen. Unter Stressbedingungen müssen letztere ihren Cortisolspiegel erhöhen
(über erhöhte ACTH-Sekretion), haben sie diese erreicht, fällt die ACTH-Sekretion via
Rückkoppelung rapide ab. Zum Ende dieser Phase haben wir bei dem ersten
Experiment das Blut bei den betäubten Individuum entnommen, was die niedrigere
ACTH-Sekretion der Hochrangigen bei dem ersten Experiment hinreichend
erklärt.
Darüber hinaus fanden wir generell bei dem Vergleich der betäubten und
unbetäubten Individuen, dass die Betäubung vermutlich erheblichen Einfluss auf die
Adrenalin-Sekretion hat, die Betäubung reduziert - nach unseren Daten ([288]) -
offensichtlich die Adrenalin-Sekretion erheblich.
Zum Zeitpunkt der Experimente lebten in unserer Kolonie auch die Mütter unserer
Alpha-Männchen, Mecki und Alba, diese testeten wir ebenfalls, beide jeweils über eine
Stunde, Blutproben entnahmen wir 5, 10, 20, 30, 40, 50 und 60 Minuten nach Betreten des
Ganges.101
Die Cortisol-Sekretion war bei Mecki zu Beginn niedriger als bei Alba, die
ACTH-Sekretion nahm bei der zweiten Probe (10 min) rapide zu und klang dann (ab
der vierten Probe (30 Minuten) sukzessive ab. Zum möglichen Einfluss des aktuellen
Ranges belegte Mecki durchgängig eine höhere Adrenalin und Noradrenalin-Sekretion
als Alba. Die Daten beider Weibchen schlossen also nahtlos an die Werte der
unbetäubten Javanermakaken an.
Wir hatten durch unsere vorläufigen Resultate ([288]) die Basis für zukünftige
Forschungsvorhaben geschaffen und in der Primatenstation auch die teure apparative
Ausstattung für die Aufbereitung der Proben installiert. Zu den geplanten
Folgeuntersuchungen (an handaufgezogenen Individuen) und Absicherungen der
Ergebnisse ist es aber aufgrund der Schließung der Primatenstation nicht mehr
gekommen.
In Unterkapitel 14.11 habe ich belegt, dass Javanermakaken leicht zu trainieren
sind, einen Zwangskäfig aufzusuchen. Die einzige Belohnung, die unsere
Makaken erhielten, war der Zugang zu den Außengehegen. Die von mir
ursprünglich verfolgte Idee, die Makaken durch Futtergaben in den Tunnel
zu locken, funktionierte nicht, alle Makaken lehnten Futtergaben im Gang
ab.102
Wir konnten selbst Blutentnahmen problemlos bei in der Gruppe
lebenden Tieren vornehmen und mussten diese nicht mehr
betäuben.103
Die Makaken merkten sich unser Vorgehen. Einmal trainiert, beherrschten sie
dieses auch Monate oder Jahre später. Zudem suchten alle in der Gruppe
geborenen Jungtiere, die das Training am Bauch ihrer Mutter erfahren haben, den
Gang auch als selbstständige Jungtiere, als subadulte und adulte Individuen
freiwillig auf. Wir hatten also gezeigt, dass das Leben in größeren Sozialgruppen
keineswegs individuelle Applikationen verhindert, dies schien uns ein wichtiger
Meilenstein auch für die gesellige Haltung von typischen Versuchsaffen zu
sein.
Doch scheidet die gesellige Haltung von Makaken für alle Versuchsvorhaben aus, bei
denen die Versuchstiere kontrolliert bestimmte Dosen eines Medikamentes zu sich
nehmen müssen, das hierarchische Sozialsystem verhindert, dass alle Individuen die
richtige Dosierung bekommen. Auch generell ist die orale Gabe von Medikamenten
schwierig, was jeder Halter von Tieren nur zu gut erfährt, noch schwieriger als bei
dem eigenen Haustier (Hund oder Katze) ist dies bei Makaken, bewahren sie
doch angebotene Tabletten in ihren Backentaschen (s. o.) auf und spucken sie
möglicherweise später wieder aus (s. u.). Insofern stellt die orale Applikation von
Medikamenten bei Makaken für den Experimentator sich als schwierige Aufgabe
dar.
Eigentlicher Auslöser für unser Forschungsanliegen war ein Fernsehbericht zur
Aufdeckung von angeblichen „Untaten“ bei Tierversuchen. Lachende Forscher hielten
an einer Kette mit einem Halsband gesicherte Javanermakakenkinder fest, fixierten den
Kopf des Versuchsaffen mit den Händen und führten in den offenen Mund eine Pistole
ein. Sie schossen sozusagen die Pille in den Rachenraum. Auf ihr Vorgehen waren die
Experimentatoren berechtigt stolz und demonstrierten es den Filmemachern. Sie
hatten aber nicht bedacht, wie Bilder auch harmloser Versuchsvorhaben wirken
können.
Gemeinsam mit meiner Doktorandin Annette Klaiber-Schuh wollte ich daher nach
einer Alternative bei Javanermakaken suchen ([114]). In mehreren Kapiteln dieses
Buches habe ich berichtet, dass unsere Affen am Morgen einen leckeren Brei erhalten,
der die Vitamin- und Eiweißbedürfnisse unserer südamerikanischen Affen zuverlässig
abdeckt. Bei den Javanermakaken war diese Gabe nicht notwendig, sie lebten gesund
ohne zusätzlichen Vitamingaben. 1995 boten wir auch unseren Makaken (Majo- und
Theo-Gruppe) Brei durch das Gitter an, alle Inividuen waren begeistert, sie stürzten
sofort an das Gitter und aßen und leckten den Brei restelos auf. Innerhalb eines
Versuchstages „lernten“ unsere Makaken in wenigen Minuten, dass der Brei lecker ist
und warteten an den folgenden Tagen bereits auf uns am Vordergitter ihres
Geheges.
In der Abbildung gibt die Skizze des Haltungsraumes Informationen über die
räumlichen Verhältnisse. Sechs Innengehege (A - F) und vier Außengehege (G - J)
standen uns zur Verfügung. In den Gehegen A und B lebten Javanermakaken
(Majo-Gruppe), in C und D Totenkopfaffen (Viktor-Gruppe), in E und F wiederum
Javanermakaken (Theo-Gruppe). Alle Affen hatten Zugang zu Außengehegen (H, I, J).
Der Tunnel wurde in Gehege C installiert. Beide Makakengruppen mussten bei diesem
Versuch einen Kreis beschreiten, die Majo-Gruppe musste via C/D, E/F, J, I, H in ihr
Innengehege (A/B) zurückgehen, die Theo-Gruppe via J, I, H, A/B, C/D zurück zu
E/F. Zusätzlich mussten wir die Totenkopfaffen trainieren, das Außengehege G (via I,
H) aufzusuchen, bevor wir mit unseren Versuchen beginnen konnten. Die Prozedur
scheint kompliziert zu sein, doch verstanden alle Affen das Vorgehen sofort, die
gesamte Prozedur dauerte nur wenige Minuten und wurde im Verlauf der
Untersuchung immer perfekter.
Die aktuelle Versuchsgruppe (Majo- oder Theo-Gruppe) wartete im Gehege A/B. Die
Individuen mussten einer nach dem anderen den Tunnel aufsuchen, eine definierte
Portion Futterbrei essen und durften dann als Belohnung D, E/F aufsuchen. Auf
Ruhebrettern in D sitzend, konnten sie das Verhalten ihrer Artgenossen im Tunnel
beobachten.104
Der Tunnel konnte durch die Schieber a - d in drei Abteile geteilt werden, in der
Mitte (2) ist die Zwangsvorrichtung, die bei diesem Versuch nicht benötigt
wurde. Das jeweilige Versuchstier hatte durch Schieber S2 Zugang zu 1 und 2.
Nach Betreten des Tunnels wurde Verbindungsschieber a geschlossen, der
Brei in 3 deponiert, danach hatte das jeweilige Inividuum durch Öffnen von c
Zugang zum Brei, zudem wurde b geschlossen. Dann begann das eigentliche
Experiment.
Zur Datenerhebung stoppten wir die Zeit
- vom Öffnen des Tunnels bis zum Aufsuchen des Tunnels durch eines der Individuen
(d. h. Öffnen des Schiebers a bis Schließen des Schiebers a);
- vom Öffnen des Schiebers c bis zum Öffnen des Schiebers d (d. h. die Zeit, die das
jeweilge Individuum benötigte nach dem Zugang zum Brei bis zur Aufnahme des
Breies bzw. bis zum Ende des Experimentes (s. u.));
- die alle Individuen der jeweiligen Gruppe insgesamt benötigten;
- für die gesamte Prozedur.
Das aktuelle Experiment endete für jedes Individuum nach der kompletten
Breiaufnahme oder - in dem Fall, in dem das Individuum es ablehnte, den Brei zu
essen, in der Lernphase 1 nach 10 min und in der Lernphase 2 (bzw. auch bei allen
weiteren Experimenten) nach 5 Minuten.
Nach den zwei Lernphasen (vgl. Tabelle 14.21) boten wir in zwölf Experimentalphasen
den Brei sechs verschiedenen Zusätzen an. Wir änderten den Geschmack, durch
Zusatz von Glukose (Phase 1) oder Salz (Phase 3), und die Farbe (Phase
2). Zudem fügten wir drei verschiedene Medikamente hinzu. In der Regel
dauerten die Experimente 5 Tage. Panacur (Panacur enthält Fenbendazol) boten
wir entsprechend der vorgeschlagenen Behandlungsdauer für jeweils 10 Tage
an.105
Zusätzliche Phasen mit unverändertem Breiangebot dienten als Kontrolle (Phase 6,
Phase 8, Phase 10, Phase 11) oder wurden durchgeführt, um eine andere Art der
Behandlung zu testen (Phase 7).
Trainingsphase Ohne Anbieten von irgendeiner Futterbelohnung trainierten wir die beteiligten Javanermakaken und Totenkopfaffen - wie oben beschrieben - siebzehn Tage lang. Alle Affen kooperierten, der Zeitaufwand für die gesamte Umsetzprozedur reduzierte sich kontinuierlich. Nachdem wir zur Überzeugung gelangt waren, die Prozedur ließe sich unmöglich weiter verkürzen, begannen wir mit den Lernexperimenten. Unabhängig von diesem Training boten wir den Javanermakaken nach dem Abschluss des Trainings unseren Experimentalbrei in den Innenanlagen an. Wir gewannen den Eindruck, dass alle Javanermakaken den Brei mochten und vor ihrem gewöhnlichen Futter präferierten. Hierdurch konnten wir sicher sein, dass der Brei - zumindest theoretisch - eine attraktive Belohnung sein könnte.
Lernphase 1
Während der Lernphase 1 folgten wir dem eigentlich von uns geplanten Vorgehen. Alle
Individuen sollten einer nach dem anderen den Brei im Tunnel essen, hierzu hatten sie
zehn Minuten Zeit. Danach wurden sie „entlassen“. Die „Bestrafung“ war also der
Aufenthalt im Gang, die „Belohnung“ das Freilassen. Eigentlich unerwartet löste nur
eines der Makakenweibchen die Aufgabe.
Allein für das Warten im Gang (10 Minuten pro Individuum) benötigten wir 140
Minuten, für das komplette Tainingsprogramm ca. drei Stunden. Dieser Zeitaufwand
schien uns für die nichtmenschlichen und menschlichen Primaten, die am Experiment
beteiligt waren, zu hoch zu sein, daher reduzierten wir ab der 2. Lernphase die
Aufenthaltszeit im Gang auf 5 Minuten pro Individuum.
Man könnte nun annehmen, die Makaken verstanden die Aufgabe nicht. Diese
Annahme ist aber - zumindest für einige Affen - falsch, vielmehr weigerten sie sich, zu
kooperieren.106
Sicher war der Lernerfolg auch so gering, weil alle Tiere gut gefüttert waren. Hungrige
Affen hätten möglicherweise anders reagiert. Doch war wesentliches Ziel des
Experimentes, jegliches Leid zu vermeiden. Zudem ist die Annahme erlaubt, dass
Hunger keinen positiven Effekt für gesellig lebende Primaten hat, er dürfte eher
agonistisches Verhalten hervorrufen, besonders bei hierarchisch organisierten Gruppen,
wie bei den Sozialgruppen der Javanermakaken.
Lernphase 2
Während der 2. Lernphase reduzierte sich sukzessive der Zeitaufwand für das gesamte
Versuchsvorhaben von zwei Stunden auf weniger als eine Stunde. Die 81 Versuchstage
verteilten sich auf einen Zeitraum von 6 Monaten, unterbrochen wurden unsere
Sitzungen gelegentlich über einen Zeitraum von zwei Tagen bis zu zwei Wochen.
Offensichtlich hatten diese Unterbrechungen keinen Einfluss auf den Lernerfolg, wie
aus den Abbildungen der individuellen Lernkurven, die in der diesem Unterkapitel
zugrunde liegenden Publikation ([114]) angegeben sind, leicht ersichtlich ist. In der
Majo-Gruppe hatten alle Individuen nach 27 Versuchstagen gelernt, was von ihnen
erwartet wurde. Für das vollständige Aufessen des Breies benötigten sie weniger als
eine Minute. Im offensichtlichen Gegensatz zu Majo, der nach der Lernkurve
offensichtlich uns und das Versuchsvorhaben testete, mal aß er den Brei innerhalb
weniger Sekunden, mal überhaupt nicht, hatten seine Weibchen das Vorgehen nicht
sofort verstanden, sie wollten offensichtlich aus dem Tunnel raus und entwickelten
„abergläubiges“ Verhalten. Z. B., einmal freigelassen nach Beißen in das Gitter des
Tunnels, versuchten sie nach Betreten des Ganges durch Beißen an verschiedenen
Stellen den „Schlüssel“ zum Öffnen zu finden. Sie mussten also als erstes lernen, dass
ihr Verhalten zwecklos und unnötig ist. Erst nach diesem Lernen begannen sie den
Brei zu kosten und zu essen. Von dem Tag an, an dem sie gelernt hatten, dass das
völlige Aufessen des Breies der „Schlüssel“ ist, kooperierten sie zuverlässig und
schnell.
In der Theo-Gruppe waren individuelle Unterschiede auffällig, fünf Tiere (Theo,
Christa, Cora, Sonja und Julia) zeigten den Mitgliedern der Majo-Gruppe
vergleichbares Verhalten, doch dauerte das Erreichen der Kannphase bei Cora, Sonja
und Julia länger. Die anderen fünf Weibchen wichen von diesem Lernprozess erheblich
ab. Ein Weibchen - Maria - verstand nach unseren Protokollen die Aufgabe vom ersten
Tag an. Doch ist es korrekter, zu sagen, sie musste die Prozedur nicht lernen. Zwar
zeigte sich eindeutig, dass sie irgendwann auch das Vorgehen gelernt haben muss, doch
können wir bei Maria den Zeitpunkt nicht angeben. Elke und Jutta hatten
offensichtlich große Probleme, die Aufgabe zu lernen. Sie kooperierten mehr oder
weniger zufällig und benötigten 60 bis 80 Tage (Lernphase 2), um die Kannphase zu
erreichen. Die letzten beiden Weibchen, Sophie und Ilse, lernten die Aufgabe nie
richtig. Dabei aß Sophie zwar die gesamte Portion auf. Nach Betreten des
Tunnels griff sie in der Regel den Brei sofort mit einer Hand, wartete dann mehr
oder weniger bewegungslos die fünf Minuten ab, nahm den Brei nach dem
Freilassen mit sich und aß ihn geruhsam auf. Bewegungslos im Tunnel zu
warten, war auch typisch für Ilse, doch berührte Ilse noch nicht einmal den
Brei.107
Die Unterschiede zwischen beiden Gruppen waren offensichtlich, das Training der
Individuen der Majo-Gruppe war nach 27 Tagen (Lernphase 2) abgeschlossen, der
Trainingserfolg bei der Theo-Gruppe blieb unbefriedigend bis zum Ende der Lernphase
2. Doch nach dem Erreichen der Kannphase durch Julia am 76. Trainingstag
verzichteten wir nach fünf weiteren Trainingstagen auf unser Ziel, restlos alle
Makaken von der Kooperationsnotwendigkeit zu überzeugen, und begannen die 12
Experimentalphasen (Tabelle 14.21).
Experimentalphasen
Zur Überprüfung des Einflusses unserer Experimente wählten wir die letzen 10 Trainingstage
der Lernphase 2 als erste Kontrolle. Offensichtlich hemmte nur Metronidazol,
dieses Medikament hat einen extrem bitteren Beigeschmack, die Akzeptanz des
Breies.108
Dieses Ergebnis veranlasste uns, in der Experimentalphase 7 die Akzeptanz einer in
dem Brei verborgenen Kapsel zu testen. Dieses Experiment schlug fehl. Am dritten
und letzten Trainingstag der Kapselphase hatten fast alle Makaken gelernt,
den Brei vollständig zu essen, die Kapsel aber entweder vor oder nach dem
Freilassen auszuspucken. Einige Tiere aßen dann die ausgespuckte Kapsel
zusätzlich auf. Kein Individuum nahm diese mit dem Brei auf, wie von uns
erhofft.
Insgesamt benötigten wir für die Theo-Gruppe viel mehr Zeit als für die
Majo-Gruppe, wobei dieser Unterschied vor allem auf das Verhalten der jeweiligen
Alpha-Männchen zurückgeführt werden kann (s. u.). Vernachlässigen wir den
Zeitaufwand für die gesamte Prozedur und bertrachten nur die Zeit im Gang,
benötigte ein Individuum im Mittel etwa eine Minute, um die Aufgabe zu lösen.
Differenzieren wir nach den Gruppen, benötigten die Individuen der Majo-Gruppe nur
wenige Sekunden, die Individuen der Theo-Gruppe mehr als eine Minute, um den Brei
aufzuessen.
Lernen durch Beobachten Wie bereits erwähnt konnten die Makaken ihre Artgenossen im Tunnel beobachten. Abhängige Kinder betraten den Tunnel am Bauch ihrer Mutter. Weder das Beobachten der perfekten Lösung der Aufgabe noch die Erfahrung, dass die eigene Mutter, den Brei gegessen hat, um den Tunnel zu öffnen, hatte einen Einfluss auf die Lernleistung des beobachtenden oder passiv teilnehmenden Individuums. Jeder Makake musste die Prozedur selber lernen. Das Aufsuchen des Ganges hingegen wurde von den passiv sich beteiligenden Jungtieren „mitgelernt“, sie hatten bei zunehmender Selbständigkeit keine Probleme, den Gang allein aufzusuchen. Zum Lernen durch Imitation waren die Javanermakaken nicht in der Lage.
Freiwillige Ausbildung einer Reihenfolge Ein weiteres wichtiges Ergebnis war
das Faktum, dass die Makaken eine mehr oder weniger feste Reihenfolge bei dem
Aufsuchen des Tunnels entwickelten. Dieses Phänomen erschwerte (Theo-Gruppe) oder
erleichterte (Majo-Gruppe) die Durchführung unserer Experimente. In der
Majo-Gruppe startete Majo (1) das Aufsuchen des Ganges, gefolgt von Laura (2),
Meike (3), Charlotte (4) und Hanne (5). Nachdem das Alpha-Männchen den Tunnel
aufgesucht hatte, waren seine Weibchen hoch motiviert, ihm zu folgen. Probleme
traten nur auf, wenn eines der Weibchen vor Majo den Tunnel aufsuchte. In solchen
wenigen Fällen versuchte Majo aktiv, den verschlossenen Tunnel aufzusuchen, das
eingeschlossene Weibchen wirkte nervös und braucht längere Zeit, um den Brei zu
essen.
Weitaus komplizierter gestaltete sich das Aufsuchen des Tunnels in der Theo-Gruppe.
Normalerweise starteten Christa oder Sophie ohne Probleme. Beide wollten den Tunnel
als erste aufsuchen, besonders motiviert war Sophie, unabhängig von der Tatsache,
dass sie die Aufgabe nicht löste oder nicht lösen konnte (s. o.). Sophie suchte aber den
Tunnel stets langsam auf, so dass andere Weibchen (Christa oder Sonja) sie
„überholen“ konnten. Nach Christa und Sophie folgten Sonja, Elke und Maria in
wechselnder Reihenfolge. Danach stoppte das Prozedere bis Theo den Tunnel
aufgesucht hatte. Als Regel suchte keines der weiteren Weibchen den Tunnel vor Theo
auf. Theo jedoch beschränkte sich häufig auf das Sitzen im Schieber, das Hineinsehen
in den Tunnel und suchte erst einmal ein Ruhebrett auf. Manchmal benötigte er allein
für das Betreten des Tunnels bis zu zehn Minuten. Nach ihm folgten Julia
und Jutta sehr schnell. Cora und Ilse waren immer die letzten Mitglieder der
Theo-Gruppe, die den Gang betraten. Die Tatsache, ob die jeweiligen Individuen die
Aufgabe gelernt hatten oder nicht, hatte offensichtlich keinerlei Einfluss auf diese
Reihenfolge.
Wie bereits in Unterkapitel 14.11 aufgezeigt, kooperieren Javanermakaken
sehr gut bei dem Trainieren, sie scheinen sogar das Training zu schätzen,
zumindest nicht als Belastung zu empfinden. Es mag unerwartet sein, dass
unsere Makaken bereitwilliger bei invasiven Forschungsvorhaben kooperierten
als bei den in diesem Unterkapitel beschriebenen nichtinvasiven Versuchen.
Vergegenwärtigen wir uns die Probleme des Individuums im Tunnel, wird dieses
Ergebnis erklärbar. Unsere Makaken wollten den Tunnel wieder verlassen (und
ihr vertrautes Gehege aufsuchen), sie waren also nicht motiviert zu essen,
selbst dann nicht, wenn das angebotene Essen eigentlich präferiert war. Die
Situation in unserem Experiment, die Makaken werden in einen kleinen Käfig
gesperrt und nichts passiert, ist sicherlich stressiger als unser invasives Handling
(Unterkapitel 14.11). Als erstes muss jedes Individuum lernen, dass es keine
Belastung ist, eine bestimmte Zeit in diesem Käfig zu bleiben, und es muss
lernen, dass es keine Chance gibt, sich diesem Aufenthalt aktiv zu entziehen.
Nach diesen „Lernerfolgen“ beginnen sie mehr oder weniger zufällig, den
angebotenen Brei zu kosten und auch zu essen. In dieser Phase - zu Beginn des
Versuchsvorhabens - machten wir Experimentatoren einen Fehler, wir belohnten schon
das Berühren des Breies durch Freilassen. Als Konsequenz aus diesem Fehler,
begannen die belohnten Individuen irgendetwas mit dem Brei zu machen
(außer Essen), z. B. verteilten sie diesen an den Wänden des Tunnels. Wir
lernten aus diesen Beobachtungen, dass es besser ist, nur die vollständige
Nahrungsaufnahme zu belohnen. In dem Moment, in dem sie die Aufgabe
verstanden hatten, begannen die Tiere sofort nach Betreten des Tunnels zu
essen.
Unsere Versuchstiere mussten drei Dinge in Folge lernen. (1) Der Aufenthalt im Tunnel
ist ohne Belastung, (2) es ist erlaubt, im Tunnel zu essen, und (3) nach dem Aufessen
des Breies erfolgt die Freilassung. Hätten wir unser Experiment mit hungrigen
Versuchstieren durchgeführt, wäre das Erreichen der Kannphase sicherlich schneller
erfolgt.109
Nicht unerwartet war für uns, dass die Makaken eine Reihenfolge beim Training
einhielten, unerwartet war aber die strikte Einhaltung dieser Reihenfolge. Für
jedes Individuum war der eigene Platz in dieser Reihe von Bedeutung. Diese
Reihenfolge war unabhängig von dem aktuellen Rangplatz, vom Geschlecht und vom
Alter110.
Ohne jeden Zweifel belegten wir, dass die Javanermakaken in unserer Kolonie
freiwillig anderen Tieren erlaubten, vor ihnen den Tunnel zu betreten, bzw.
dass sie nach erst nach Betreten durch bestimmte andere Individuen diesen
folgten.111

1Javanermakaken werden von Altmeister Brehm ([12]) nicht erwähnt, er berichtet über den „gemeinen Makaken“, identifiziert diesen aber als Hutaffe, was wohl ein Fehler ist, nach Hill ([88]) war nämlich „der gemeine Makak“ der deutsche Name für unseren Javanermakaken. Nach Heck ([68]) ist er der Makake schlechthin. „Die Art, die der ganzen Gruppe den Namen gegeben hat, ist der langschwänzige Makak oder Javaneraffe ... “([68], Seite 533).
2In Indien verehren die Hindus den Affengott Hanuman, daher sind seine Nachfahren, die Hanuman-Languren Presbytes entellus, besonders geschützt. Da nun die normalen Inder - wie die normalen Deutschen - zwischen den verschiedenen Affenarten nicht differenzieren können (Affe = Affe) genießen auch die Rhesusaffen religiösen Status. Gegen das Abfangen entstehen so lokale Proteste, was die Indische Regierung bewogen hatte, den Export zu unterbinden.
3Hätte Harry F. Harlow seine berühmten Deprivationsexperimente an Indischen Hutaffen durchgeführt, hätte er den Einfluss der Deprivation auf das Verhalten nicht belegen können. Seine Experimente, die in Nordamerika und Europa zur segensreichen Änderung des Umgangs mit Waisenkindern geführt haben, wären für uns Menschen folgenlos geblieben.
4Meine Mitarbeiter und ich mochten auch unsere Makaken, nach der Geburt von Frieda (30.01.1975) und Paula (10.03.1975) demonstrierten sie täglich die sprichwörtliche „Affenliebe“ bei der Jungenaufzucht. Wir registrierten zudem ihr auffälliges Dominanzverhalten, doch investierten wir absichtlich keine Zeit in das Beobachten der Tiere. Zum einen war das Verhalten der Makaken nach meinem damaligen Wissensstand umfänglich beschrieben und bekannt, zum anderen entwickeln Beobachter enge emotionale Beziehungen zu ihren Tieren, diese wollten wir nicht entstehen lassen, waren doch die Makaken für anatomische und experimentelle Forschungsanliegen bestimmt.
5Den Lesern meines Buches wird auffallen, dass die Qualität der Arbeit an Javanermakaken sich von der an den bisher besprochenen Arten unterscheidet. Bei den anderen Arten bemühten wir uns, durch unsere Versuchsvorhaben möglichst wenig in das Sozialleben einzugreifen. Wir wollten grundlegende Fragen der Haltung klären (Haltungsexperimente) und versuchen, Zuchtkolonien aufzubauen. Bei den Javanermakaken hingegen führten wir gezielte Experimente zur Klärung noch offener Forschungsfragen durch.
6Klaiber, A.: Der Einfluß der Gruppenstruktur und der Käfiggröße auf das Verhalten des Individuums bei dem Javanermakaken Macaca fascicularis, 1994.
7Engelhardt, R.: Die sozialen Interaktionen im 6. Lebensmonat bei handaufgezogenen Javanermakaken Macaca fascicularis. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe und die Oberstufe. Kassel, 29. April 1985.
Erbarth-Fischer, M.: Die Entwicklung handaufgezogener Javanermakaken Macaca fascicularis während des ersten halben Lebensjahres unter besonderer Berücksichtigung der Entstehung des sozialen Beziehungsgefüges. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe und die Oberstufe. Kassel, 14. Oktober 1985.
Fiege, C.: Das Verhalten von in einer „peer“-Gruppe aufgezogenen Javanermakaken Macaca fascicularis während des dritten Lebenshalbjahres. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe und die Oberstufe. Kassel, 23. September 1986.
Sobisch, E.: Zum Präferenzverhalten von „peer“-aufgezogenen Javanermakaken Macaca fascicularis. Wissenschaftliche Hausarbeit zur ersten Staatsprüfung für das Lehramt für die Mittelstufe und die Oberstufe. Kassel, April 1987.
8Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1985 publizierten Beitrag ([246]).
9Heute bin ich überzeugt, dass es Freundschaften bei Makaken überhaupt nicht gibt, die Freundschaften sind vielmehr strategische Beziehungen zu dem theoretisch und praktisch für das Individuum gefährlichsten Sozialpartner, dem Rangnachbar. Dieser muss kontrolliert werden, am einfachsten durch enge Sozialkontakte (Kontaktsitzen und Beziehungen gegenseitiger sozialer Körperpflege). Mehrfach musste ich über die Jahre beobachten, dass zwei über viele Monate scheinbar enge „Freundinnen“ plötzlich und für uns unerwartet sich massiv bekämpften. Kurz nach dem Kampf waren die engen Beziehungen zwischen den „Freundinnen“ wieder erneut monate- oder jahrelang beobachtbar, doch hatte zwischen den Individuen ein Rangwechsel stattgefunden.
10Ein unbeabsichtigtes Ergebnis war, dass alle Makaken-Kinder sich bevorzugt in dem engen Verbindungsgang dichtgedrängt aufhielten. Der Gang war der höchste Ort der Anlage (auch alle anderen Makaken in der normalen Haltung bevorzugten höher angebrachte Ruhebretter), zudem wohl auch der interessanteste, von hieraus konnten sie jeden eintretenden Besucher und mich und meine Mitarbeiter am Schreibtisch beobachten.
11Während der jahrelangen Aufzucht wechselten auch die studentischen Mitarbeiter. Unsere Makakenkinder differenzierten zwischen Studenten, die vor ihnen präsent waren und solchen, die nach ihnen das Versuchsvorhaben begleiteten, jene wurden als ranghöher behandelt, diese als rangniedriger. So konnte es meinen engagierten Mitarbeitern passieren, dass sie bei Eintritt „ranghöherer“ Mitarbeiter plötzlich von den kleinen Makakenkindern angegriffen und gebissen wurden. Ich hatte das Glück, dass mich mit Ausnahme von Laura, die offensichtlich meine Mitarbeiterin Cornelia Schäfer-Witt als ranghöher einschätzte, alle Makakenkinder als „ranghöher“ einstuften, ich wurde also von Bißattacken verschont.
12Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1980 publizierten Beitrag ([275]).
13Für die Haltung und Beobachtung der Tiere standen fünf Innenkäfige zur Verfügung. Zu Beginn der Versuche waren Nikita, Vanda und Vera in Käfig 1 (1,20 m x 3,00 m x 2,50 m), Vema, Vau, Blonda, Jungfrau und Zita in Käfig 2 (1,20 m x 1,80 m x 2,50 m) und Alba, Dua, Stirni, Mecki und Omega in den Käfigen 3 - 5 (1,20 m x 1,00 m x 2,50 m) untergebracht. Nach Fusion aller Tiere dienten die Käfige 1 und 2 ausschließlich zur Haltung der Gesamtgruppe, während die Käfige 3 - 5 auch zur vorübergehenden Separierung einzelner Tiere genutzt wurden. Alle Käfige waren durch Schieber (50 cm x 50 cm) miteinander zu verbinden. Seit Oktober 1975 standen den Tieren noch zusätzlich ein Außengehege zur Verfügung. Die Käfigeinrichtung besteht aus Holzbrettern und Holzleitern. In den Haltungsraum fiel Tageslicht, zusätzlich wurde er täglich von 7.00 - 19.00 Uhr über eine Schaltuhr mit Neonlampen beleuchtet. Der Haltungsraum war vollklimatisiert, seine Temperatur schwankte dabei systematisch von 23 Grad C - 27 Grad C (relative Luftfeuchtigkeit 60 - 70 %). Das Außengehege stand den Tieren unabhängig von der Außentemperatur von 8.00 - 19.00 Uhr zur Verfügung.
14Aus den Beobachtungen ergab sich die Dominanz, z. B. (1) A und B sitzen auf einem Brett, A holt sich das Futter, geht zurück auf das Brett und frisst, B bleibt sitzen, d. h. A ist dominanter als B; (2) A und B sitzen auf dem Brett, A holt sich das Futter, flieht oder bleibt auf dem Boden sitzen, B droht bzw. verfolgt A, d. h. B ist dominanter als A.
15Dies korrespondiert Kummers „Regel 4“: „A dyad can be reduced to lower stages under the influence of other conspecifics....“ ([117], Seite 136).
16Er bedrohte diejenigen Weibchen, die Anstalten machten, ein im Hintergrund des Käfigs angebrachtes Ruhebrett zu verlassen.
17Dabei bedrohte er auch gelegentlich das neue Weibchen bedrohend, bzw. griff nach ihm.
18Vera war einer der Versuchsaffen von Werner Meinel. Eigentlich sollte sie perfundiert werden, doch konnte ich Werner Meinel überzeugen, dass für sein Experimentalziel, das Einwachsen künstlicher Zahnwurzeln zu untersuchen, das Töten von Vera keine zwingende Voraussetzung war, eine Teilamputation des Unterkiefers lieferte hinreichende Resultate.
19Diese Alpha-Position behielt Vanda dann über mehr als ein Jahrzehnt unangefochten (vgl. Unterkapitel 14.9).
20Dann stürmen die übrigen Gruppenmitglieder herbei und packen sich schnellstmöglich beide Backentaschen - ein Merkmal des Genus Macaca - voll und fliehen auf eines der oberen Ruhebretter, um in Ruhe zu essen. In den Backentaschen ist die Nahrung in der Regel sicher aufbewahrt. Nur Vanda stahl regelmäßig Futter aus dem Mund der Sozialpartner. Die Backentaschen werden von Jungtieren auch genutzt, um vermeintlich Wertvolles - wie z. B. Glasscherben - zu bewahren. Dann hat der Halter oft tagelang Sorgen, bis die Jungtiere irgenwann ihren „Besitz“ aufgeben.
21Das auf den Beobachter sehr gefährlich wirkende spielerische Beißen ist wahrscheinlich auch ein Vertrautheitstest, würde das Jungtier dem Biß entfliehen, wären erhebliche Verletzungen unvermeidlich.
22Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1982 publizierten Beitrag ([273]).
23Die Diskussion, ob in Primatengesellschaften überhaupt eine Rangordnung vorhanden ist oder ob die einzelnen Tiere nur bestimmte Rollen wahrnehmen, soll hier nicht erneut geführt werden, hierauf bin ich bereits eingegangen (vgl. 14.1).
24Wir können zwar nicht ausschließen, dass unsere Bedingungen in Menschenobhut die Ergebnisse beeinflussen, doch gibt es die unbeeinflussten Affengruppen nicht, so sind z. B. Gruppengrößen auch im Freiland oft durch Abfangen, Zufütterung u. ä. „human-dependent“. Über den Einfluss der Käfiggröße liegen unterschiedliche Angaben vor, Untersuchungen geben Hinweis darauf, dass ein engerer Raum sogar zur Reduktion agonistischen Verhaltens führen kann, „crowding“ Effekte sind also kaum nachweisbar. In jeder beobachteten Gruppe im Freiland und Labor bleiben jedoch die zwei grundsätzlichen und auch für unsere Untersuchung bestimmenden Komponenten der Makakensozietät, nämlich die Dominanzhierarchie und die „kinship relations“ unverändert erhalten, so dass bei Aussagen hierzu der Haltungseffekt vernachlässigt werden kann.
25Zusätzlich wurden die Interaktionen der Gruppe (dreißig Minuten) und die Interaktionen von zwei Einzelindividuen (jeweils fünfzehn Minuten) über eine Video-Kamera aufgezeichnet. Bei dem während unserer Experimente notwendigen gleichzeitigen Protokollieren von zwei Gruppen wurde auf das Beobachten der Einzeltiere verzichtet, stattdessen protokollierten wir die Interaktionen beider Gruppen jeweils zwei Stunden lang. Abweichend von diesem generellen Versuchsplan verzichteten wir bei den Versuchsabschnitten E.S. 3.4 bis 3.8 auf das Verschieben der Beobachtungszeit, hier fanden die Beobachtungen stets zwischen 15.00 bis 18.00 Uhr statt.
26Entscheidend änderte sich nur die soziale Position des Weibchens Mecki; diese erkämpfte sich von Juli 1976 bis Februar 1977 sukzessive den vierten Rangplatz in der Weibchenhierarchie. Strategisch nutzte sie die agonistischen Interaktionen der Alpha-Tiere aus, drohte mit diesen, biss dann die fliehenden oder „pressenden“ Tiere und erlangte, hierdurch einen höheren Rang. Mecki selber blieb dabei nicht unverletzt, zahlreiche vernarbte Platzwunden waren Zeichen dafür, dass ihr Aufstieg nicht ohne Gegenreaktion der angegriffenen Tiere erfolgte. Bereits im Januar 1977 konnten wir zudem beobachten, dass Mecki - im Zusammenhang mit gegen Vera gerichtetem agonistischen Verhalten des Alpha-Männchens - Vera gleichfalls angriff, doch konnte diese sich hier behaupten und 1977 durchgängig ihre höhere Position gegenüber Mecki halten: Erst im Mai 1978 - eventuell im Zusammenhang mit dem Verlust von Veras ältester Tochter Paula (25. 04. 1978) - erreichte Mecki die dritthöchste Rangposition innerhalb der Weibchenhierarchie. Weitere Rangänderungen in der Gruppe beschränkten sich auf minimale Verschiebungen zwischen rangbenachbarten Individuen. Blonda tauschte den Rangplatz mit Zita (und erreichte somit wieder die Position, die sie bereits einmal (bis September 1978) in der Weibchenhierarchie eingenommen hatte. Darüber hinaus gelang es Jungfrau im Frühjahr 1978, den Rangplatz mit Stirni zu tauschen.
27So bedrohte Majo bei der ersten (E.S. 1.1) und zweiten (E.S. 1.7 ff.) Separation Nikitas Orbi, bei der zweiten Separation zudem auch Berni; Tritus: Ernst und Paul (E.S. 1.7); Ernst: Orbi(E.S. 1.7); Orbi: Max (E.S. 1.9) und Protus (E.S. 1.7); Max: Protus und Paul.
28So drohte Majo: Calva (E.S. 1.9), Vera (E.S. 1.7 und 1.9), Barbara (E.S. 1.7 ff..), Alba (E.S. 1.9), Jungfrau (E.S 1.8), Christa (E.S. 1.8); Tritus: Barbara (E.S. 1.7), Jungfrau (E.S. 1.1 und 1.7); Berni: Flava (E.S. 1.1 und 1.7), Itta (E.S. 1.9); Orbi: Alba (E.S. 1.7 ff.), Jungfrau (E.S. 1.1 und 1.7), Itta (E.S. 1.1), Omega (E.S. 1.8); Max: Itta (E.S. 1.1 und 1.9), Christa (E.S. 1.8), Omega (E.S. 1.9) ; Protus: Itta (E.S. 1.8).
29Es sei jedoch bemerkt, dass Zita es bei Introduktion von Vera schaffte, durch gemeinsames Drohen mit den „künstlich Ranghohen“ die dritthöchste Position in der Gruppe nach Stirnis Tochter Itta zu erhalten. Wohl durch den Aufbau besonders enger Kontakte zu dem ranghöchsten Weibchen Stirni bereits in den ersten Tagen nach Introduktion (E.S. 2.5 ff.) erreichte sie in den vier Wochen gemeinsamer Haltung eine bessere Ausgangsposition und war außer an agonistischen Interaktionen gegenüber den später Introduzierten, besonders gegenüber Vera, nun auch an solchen gegen Jungfrau, Vau und Omega beteiligt (E.S. 2.7); zudem sei darauf hingewiesen, dass sich Vera im Gegensatz zu Barbara und Blonda nach Introduktion auch gegenüber Jungfrau durchsetzte, also mit Ausnahme von Zita gegenüber allen vor ihr in die Ausgangsgruppe introduzierten Weibchen.
30Auffallend bei den Introduktionen war das hohe agonistische Potential der rangmittleren Weibchen und ihrer Kinder. Sie drohten nicht nur den Neuintroduzierten, sondern näherten sich den widerstandslos auf dem Boden pressenden oder am Gitter hängenden Individuen, bissen diese in den Rücken, in die Arme und Augenbrauen; Verhaltensweisen, die wir sonst in der Gruppe nicht beobachten können. Es war zudem auffällig, dass adulte Weibchen adulte Weibchen, juvenile Männchen juvenile Männchen, juvenile Weibchen juvenile Weibchen und Infantes Infantes bedrohten, verfolgten und bissen. Allein Jungfrau war im Verlauf des Versuches in der Lage, sich gegenüber Stirni, dem rangniedrigsten und am stärksten bedrohtesten Weibchen dieser Gruppe, ihrem alten Rangnachbarn in der Gesamtgruppe, durchzusetzen. Die später introduzierten Omega und Vau zeigten auffällig auch kein Rangsicherungsverhalten, wie wir es für die Rangmittleren beschrieben haben.
31Es dauerte einige Tage, bis Mecki und ihre Töchter sich wieder frei bewegen konnten.
32Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1982 publizierten Beitrag ([274].
33„it even ’made things worse’ “ ([34], Seite 578.)
34In Unterkapitel 14.9 werde ich belegen, dass auch die Qualität des Alpha-Männchens von hoher Bedeutung ist.
35Dabei war ihr Interesse stets auf die Mutter eines bestimmten Jungtieren gerichtet. Es bestand dabei eine klare Tendenz, zu weiblichen Kindern länger Kontakt zu halten (Zita, Mecki), was aber nicht hinreichend gesichert ist, da nämlich gerade die beiden letzten Jungtiere weiblich waren, es also auch sein kann, dass sie - wenn jüngere Kinder nicht vorhanden sind - Kindern unabhängig vom Geschlecht auch über das Alter von 60 Tagen hinaus hohe Aufmerksamkeit schenkt.
36Neben der grundsätzlich herausragenden Position des Alpha Weibchens fiel besonders dessen enge Beziehung zum Alpha-Männchen auf. Wie wir aufgezeigt haben, ist das Alpha-Weibchen jedoch eine „Institution“, wie das Alpha-Männchen, offensichtlich unabdingbar für das Funktionieren einer Makakensozietät.
37Das besondere Verhalten unseres Beta Weibchens Frieda könnte theoretisch auch damit verbunden sein, dass Frieda in einer Sondersituation aufwuchs. Als erstes in Kassel geborenen Jungtier wurde sie, bedingt durch die soziale Stellung der Mutter, von Geburt an als hochrangig behandelt und wuchs ohne Kontakt zu anderen Jungtieren, also ohne altersgemäße Spielkontakte auf. Mit der frühkindlichen sozialen Situation wäre eventuell das besondere Verhalten hinreichend erklärt. Betrachten wir aber nun unsere Versuchsergebnisse und hier die Verhaltensweisen der Töchter der jeweiligen ranghöchsten adulten Weibchen, dann scheint es richtiger zu sein, Friedas Verha!ten als das Verhalten der Tochter des ranghöchsten Weibchens richtig zu umschreiben. Es spricht vieles dafür, dass die Tochter des Alpha-Weibchens ebenso eine Institution ist wie das Alpha-Männchen und das Alpha-Weibchen und dass sie mit diesen gemeinsam das Zusammenleben in Makakengruppen reguliert. Dies gilt natürlich nur, wenn das Alpha-Weibchen auch weibliche Nachkommen hat. Es könnte vermutet werden, dass langfristig das Halten einer Alpha-Position nur bei Vorhandensein einer Tochter und damit eines verlässlichen langfristigen Koalitionspartners möglich ist.
38Während eines Oestrus reduzieren adulte Weibchen ihre Kontakte zu anderen Gruppenmitgliedern, inklusive ihren eigenen Kindern. Da das Besteigen durch das Alpha-Männchen auch außerhalb des sexuellen Kontextes als Besitzdemonstration u. ä. erfolgt, können wir hier nur „erfolgreiche“ Oestren berücksichtigen, also Oestren, die auch später zur Geburt eines Kindes führten. Während des Beobachtungszeitraumes fielen diese Oestren in drei zeitlich getrennte Perioden, nämlich 1. Omega (E.S. 1.5); 2. Jungfrau (E.S. 2.11), Vera (E.S. 3.0), Stirni (E.S. 3.1); 3. Blonda/Zita/Barbara/Alba (E.S. 5.1 und 5.2).
39Theoretisch könnten 3 verschiedene Grundgesetzmäßigkeiten für die Präferenz juveniler Männchen verantwortlich sein, nämlich: 1. juvenile Männchen halten vornehmlich Kontakte zu juvenilen Männchen, zu deren Müttern ihre eigenen Mütter enge soziale Beziehungen unterhalten; 2. juvenile Männchen unterhalten vornehmlich Kontakte zu juvenilen Männchen der gleichen Altersstufe; und 3. juvenile Männchen unterhalten vornehmlich Kontakte zu juvenilen Männchen, deren Mütter ihren eigenen Müttern rangbenachbart sind. Überprüfen wir diese 3 Hypothesen an unseren Daten, so zeigt sich, dass nur mit der 3. Hypothese die Beziehungen der juvenilen Männchen untereinander hinreichend erklärt werden können. So hat Majo vornehmlich Kontakt zu Tritus, Berni, Ernst, Orbi; Tritus zu Majo, Berni, Ernst, und nach dem ersten Rangabfall von Vera (E.S. 2.7 ff.) auch zu Orbi, bzw. nach dem zweiten Rangabfall von Vera (E.S. 3.5 ff.) zusätzlich zu Max und Paul. Berni und Ernst unterhalten vor allem Kontakte zu Majo, Tritus, Orbi und Max, wobei - berücksichtigen wir nicht die Geschwister-Beziehungen zwischen Berni und Ernst - die zu Orbi die intensivsten sind, Orbi schließ1ich zu Majo, Berni, Ernst und Max, wobei die Kontakte zu Berni am intensivsten sind. Die Mütter der jeweiligen Männchen sind über Tabelle 14.2 zugänglich.
40So hat Calva vornehmlich Kontakt zu Flava, der Tochter der rangnächsten Tochter-Mutter und seltener zu Itta (vornehmlich nach Separierung von Zita und Flava, wo Itta als einziges juveniles Weibchen noch in der Gruppe verblieb); auffällig wenig Kontakt unterhält sie zu Christa. Flavas Beziehungen zu anderen juvenilen Weibchen lassen entsprechend der sozialen Position der Mutter im mittleren Bereich der Hierarchie keine klaren Präferenzen für bestimmte andere Tiere erkennen. Itta bevorzugte Christa vor Flava und Calva, sehen wir einmal von den intensiven sozialen Beziehungen zu Calva bei Fehlen von Christa ab. Christa schließlich hat die engsten Beziehungen zu Itta, geringere zu Flava und die geringsten zu Calva, ihre Kontakte entsprechen also auch hier der Rangposition der Mutter.
41In jeder Gruppe nimmt ein Männchen die Rolle des Alpha-Männchens, ein Weibchen die Rolle des Alpha-Weibchens und, soweit dieses eine Tochter hat, ein Weibchen die Rolle der Tochter des Alpha Weibchens wahr. Diese drei kontrollieren alleine und gemeinsam alle Interaktionen der Gesamtgruppe. Bei den übrigen Individuen der Gruppe war auffällig, dass bei Sozialkontakten Tiere des gleichen Alters und Geschlechtes anderen vorgezogen wurden, so suchten adulte Weibchen die Nähe adulter Weibchen, juvenile Männchen diejenige juveniler Männchen und juvenile Weibchen den Kontakt zu juvenilen Weibchen.
42Darüber hinaus kommt Familienbeziehungen höchste Bedeutung zu, Kinder derselben Mutter halten untereinander und mit ihrer Mutter enge Sozialkontakte, dabei ist das Geschlecht der Geschwister ohne Bedeutung. Die Kontakte zwischen Müttern und Töchtern sind intensiver als solche zwischen Müttern und Söhnen; bei den weniger engen Beziehungen zu ihren Söhnen sind die Mütter relativ häufiger aktiver Partner als bei denjenigen zu ihren Töchtern.
43Diese Gesetzmäßigkeiten werden schließlich überlagert durch die für Macaca auffällige lineare Rangordnung. Der Rang in der Gruppe geborener Jungtiere ist abhängig vom Rang ihrer Mütter, so nehmen Weibchen - unabhängig vom Alter - in Relation zu anderen Weibchen den Rangplatz wie ihre Mütter ein; der Rang der juvenilen Männchen ist darüber hinaus auch abhängig vom Alter. In jeder Altersgruppe ist der Rang der einzelnen Individuen jedoch nach dem Rang der Mütter vorhersagbar. Hauptverantwortlich für die Stabilität der Rangstruktur ist die Tendenz aller Individuen, vor allem mit rangbenachbarten Tieren enge soziale Beziehungen aufzubauen, dies gilt für adulte Weibchen, juvenile Männchen und juvenile Weibchen. Juvenile Weibchen haben zudem auch besonders enge Beziehungen zu adulten Weibchen, zu denen ihre eigenen Mütter enge soziale Kontakte unterhalten und zu Töchter-Müttern, die rangniedriger sind als die eigene Mutter. Dieses offensichtliche „Wissen“, welche Weibchen rangniedriger sind als die eigene Mutter, ist auch bei juvenilen Männchen aufzeigbar, so bedrohen diese bei Abwesenheit des Alpha-Männchens vor allem nächstrangige Männchen und versuchen, Weibchen zu kontrollieren, die rangniedriger sind als die eigene Mutter.
44Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1982 publizierten Beitrag ([291]).
45Kontaktsitzen, Zusammensitzen, Spielen (verschiedene Formen des Spielens)
46Sichtsetzenzu, Schmatzen, Umarmen, Soziale Körperpflege, Putzaufforderung, Olfaktorischer Kontakt, Tragen eines Jungen.
47Drohfixieren, Drohen mit Grunzen, Scheinangriff, Hetzjagen, Zerren, Beißen, Bedrängen bzw. Schreien, Weichen, Fliehen, Ignorieren.
48Hängen am Bauch, Klettern am Bauch, Hängen auf Rücken, Greifen nach; An der Mutter (Zeit), An der Mutter und Zitze im Mund, Ohne Körperkontakt zur Mutter (Zeit), Annäherung von der Mutter ausgehend, Sich entfernen von der Mutter/von dem Kind, gleichzeitigem Entfernen, Unterstützung des Kindes, Festhalten des Kindes; Nicht soziales Spiel, Gegen sich selbst gerichtetes Verhalten (Sichkratzen, Sichputzen, etc.), Manipulieren.
49Für die Ermittlung der Ergebnisse addierten wir bei den Interaktionen die Daten eines Monats und bildeten einen Mittelwert (bezogen auf 15 Minuten). Für die Zeitangaben errechneten wir für jeden Lebensmonat den Prozentanteil des „An der Mutter“- bzw. „Ohne Körperkontakt zur Mutter“-seins, für den „Gesamteindruck“ den Prozentanteil der einzelnen „Eindrücke“ an der Anzahl der gesamten „Eindrücke“ eines Monates.
50Den Erwartungswerten liegt die theoretische Annahme zugrunde, dass ein bestimmtes Individuum zu jedem anderen Tier der Gruppe, bzw. der entsprechenden Altersgruppe (etc.) gleichviel Kontakte unterhält.
51Weitere individuelle Unterschiede, so z. B. die Zunahme der Zeit „An der Mutter“ im fünften Lebensmonat bei Felix und Angela bzw. im sechsten Lebensmonat bei Micha, Olga und Tina sind im Rahmen dieser Arbeit ohne Bedeutung und sollen hier nicht diskutiert werden. Es sei jedoch betont, dass eine Beziehung zum Ranggefüge der Weibchen nicht gegeben ist. Dies ist unschwer zu erkennen, wenn wir die Kinder nach den Rängen ihrer Mütter reihen (Männchen: Toto/Micha/Neo/Felix; Weibchen: Olga/Angela/Tina/Ursula). Es scheint eher so, dass spätergeborene Männchen später selbständig werden als frühergeborene. Eine entsprechende Tendenz ist bei den weiblichen Kindern nicht erkennbar, hier fällt auf, dass Töchter ranghöherer Weibchen im zweiten Lebensmonat noch häufiger ausschließlich von der Mutter getragen werden als diejenigen rangniedrigerer Mütter. Dies dürfte daran liegen, dass die entsprechenden Mütter, Barbara und Mecki, weit eher als Zita und Alba das Tragen durch die beiden ranghöchsten Weibchen (Vanda und Frieda) verhindern konnten.
52Sobald das Interesse von Vanda und Frieda nachließ, diese unaufmerksam wurden, sprangen die Mütter herbei, griffen ihr Junges und pressten es an sich. Rangniedrigere Mütter konnten dabei weniger Widerstand leisten. Insofern ist es nicht verwunderlich, dass Vanda deren Kinder präferierte.
53Berücksichtigen wir jedoch, dass weibliche Kinder für männliche Jungtiere völlig unattraktiv waren, dann finden wir auch hier, dass (relativ) männliche ältere Juvenile eindeutig mehr mit männlichen Kindern interagieren, also eine klare Bevorzugung des eigenen Geschlechts. Es muss hier jedoch betont werden, dass die Beziehungen zu zwei-, drei- und vierjährigen Juvenilen bzw. Subadulten für die heranwachsenden Kinder ohne große Bedeutung sind. Die Einjährigen hingegen interagieren sehr häufig mit den jüngeren Kindern, dabei sind weibliche Kinder nur für weibliche Juvenile attraktiv. Dies dürfte auch hinreichend erklären, warum weibliche Kinder insgesamt weniger Sozialkontakte zu dieser Altersgruppe unterhalten als männliche, waren doch nur ein Drittel der Individuen der entsprechenden Altersgruppe weiblich. Auffällig suchten diese jedoch sowohl männliche als auch weibliche Kinder stärker zu pflegen als ihre männlichen Altersgenossen. Die Pflegeappetenz ist demnach bei allen weiblichen Individuen unabhängig vom Alter in weit höherem Masse ausgeprägt als bei den entsprechenden Männchen. Neben diesem durch das Geschlecht des einzelnen Tieres mehr oder weniger vorbestimmten Interaktionsmuster fällt darüber hinaus der Kontakt zu Gleichalten besonders auf. Diese Spielkontakte sind offensichtlich für die Heranwachsenden bestimmend. Auch hier konnten klare Präferenzen für gleichgeschlechtliche Altersgenossen aufgezeigt werden. Die Deutung dieses Befundes ist schwierig, da zwangsläufig die weiblichen Kinder den männlichen nur bedingt zur Verfügung standen. Die Daten von Toto weisen darüber hinaus daraufhin, dass fast gleichalte Kinder anderen vorgezogen werden, so unterhielt Toto, wenn auch geringfügig, relativ mehr Kontakte zu weiblichen als zu männlichen Altersgenossen. Vergleichen wir jedoch seine Daten mit denen von Olga und den übrigen Weibchen, dann wird auch hier eine stärkere Präferenz der männlichen Kinder für männliche Altersgenossen deutlich.
54Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1986 publizierten Beitrag ([285]).
55Zusätzlich gaben wir jeweils den Erwartungswert an (14.5.1). Dem Erwartungswert liegt die theoretische Annahme zugrunde, dass jedes Individuum der Gruppe bzw. der entsprechenden Alters- bzw. Geschlechtsgruppe an den Interaktionen des jeweiligen Jungtieres gleichermaßen beteiligt war.
56Dabei wurde der Prozentanteil jeder Altersgruppe, bzw. des Alpha-Männchens, des Alpha-, des Beta-Weibchens und der adulten Weibchen (ohne die jeweilige Mutter) an der Gesamtzahl der positiv-sozialen Interaktionen des jeweiligen Jungtieres zu allen anderen Gruppenmitgliedern (ohne Mutter) errechnet und mit dem Erwartungswert verglichen.
57Offensichtlich wurde das Wahlverhalten gegenüber adulten Weibchen von dem Verhalten der Mütter beeinflusst. Generell fanden wir, dass alle acht Juvenilen im zweiten Lebenshalbjahr viele Kontakte zu den Weibchen unterhalten, mit denen auch die jeweilige Mutter häufig interagiert, wenige dagegen zu solchen, die für die eigene Mutter unattraktiv sind. Die genauere Analyse des Datenmaterials erbrachte, dass bei den 72 in unserer Gruppe möglichen Paarungen Kind/adultes Weibchen (Nichtmutter) in 59 Fällen (82 %) eine Übereinstimmung zwischen Wahl des Kindes und Wahl der Mutter festzustellen war (23mal unterhielten Mutter und Kind enge, 55mal keine Kontakte zu dem jeweiligen adulten Weibchen). Achtmal beobachteten wir nur geringe Kontakte zwischen Kind und adultem Weibchen, obwohl letzteres von der Mutter des Kindes präferiert wurde. Fünfmal schließlich interagierten Jungtiere mit Weibchen, die wenig mit ihrer eigenen Mutter interagierten, dabei war auffällig, dass in allen fünf Fällen das gewählte Weibchen rangniedriger war als die eigene Mutter.
58Diese generellen Ergebnisse gelten unabhängig von den auffälligen individuellen Unterschieden, die wir zwischen den acht Jungtieren beobachten konnten. Wie bereits ausführlich diskutiert, kann hierfür die unterschiedliche Erfahrung der Mütter bei der Jungenaufzucht, für die wir - nach Beobachtungen an vielen unserer Nachzuchtweibchen - zahlreiche Belege haben, nicht verantwortlich sein, da alle acht Mütter Multipara waren. Auch das Geschlecht der Juvenilen kann wegen der hohen Abweichung bei Individuen beiderlei Geschlechtes nicht für die beobachteten individuellen Unterschiede verantwortlich sein, behandelten doch die Mütter männliche und weibliche Kinder nicht unterschiedlich, sie blieben auch im zweiten Lebenshalbjahr weiterhin tolerant, offensichtlich unabhängig vom Geschlecht. Da alle männlichen Individuen vor den weiblichen geboren wurden, ist damit auch der unterschiedliche Geburtstermin kein Parameter, der für die beobachteten individuellen Unterschiede verantwortlich sein kann.
59Die Bedeutung des Alters für die Individuen der Untersuchungsgruppe wird auch bei den Kontakten zu Juvenilen anderer Altersgruppen deutlich. Alle interagieren zwar häufig mit Individuen des eigenen Jahrganges, mit den 1981 geborenen Jungtieren interagieren dagegen nur die Jungtiere, die 1980 später geboren wurden. Für die älteren drei Männchen waren die jüngeren offensichtlich unattraktiv, sie wählten stattdessen den Kontakt zu den jüngsten Individuen des vorangegangenen Jahrganges.
60Ziehen wir nämlich auch in Betracht, wie häufig die acht im Focus stehenden Tiere überhaupt mit den Individuen der anderen Altersgruppen interagiert haben, erkennen wir, dass die Anzahl der Interaktionen zur Gesamtheit der Individuen der jeweiligen Altersgruppen nur in einigen Ausnahmen (Neo zu 1979er und 1976er, Micha zu 1976er, Felix zu 1979er, Tina zu 1976er und Angela zu 1978er) überhaupt über dem nach der Anzahl der Individuen zu erwartenden „Erwartungswert“ liegt.
61Gerade im Hinblick auf Erhebungen an den ohne Einfluss von Müttern und Geschwistern aufwachsenden „peers“ können wir hier nicht entscheiden, worauf diese Unterschiede beruhen. zweifellos wäre es aber hinreichend anzunehmen, dass die jungen Makaken - bedroht durch ranghöhere Artgenossen - den Kontakt zu ihren Müttern suchten, um dort Schutz zu finden. Dafür spricht auch, dass die Tochter des ranghohen Weibchens Mecki, Angela, das Selbständigste aller Jungtiere war. Für dieses Jungtier war zudem auffällig, dass es als einziges der acht untersuchten Tiere häufig mit Alpha-Weibchen und Alpha-Männchen interagierte. Fragen wir uns, warum nur Angela noch verstärkt mit dem Alpha-Männchen und Alpha-Weibchen interagiert, dann können zwei verschiedene Gründe dafür verantwortlich sein, so ist Angela das jüngste Kind der acht untersuchten Kinder. Da jeweils die jüngsten besonders attraktiv sind, könnte dies hinreichend sein, die gezeigten Präferenzen zu erklären. Dagegen spricht aber, dass noch weitere Jungtiere geboren wurden, also Angela keineswegs jüngstes Individuum der Gruppe blieb. Suchen wir nach weiteren Kriterien, die es erlauben Angela von den übrigen sieben Jungtieren zu diskriminieren, dann scheint das einzige Kriterium der Rang der Mutter zu sein. Mecki war als einzige der vier Töchter-Mütter ranghoch und dementsprechend auch rangbenachbart mit dem Alpha-Weibchen. Dass rangbenachbarte Weibchen besonders enge Sozialkontakte unterhalten, wissen wir bereits. Mit dem Rang der Mutter wären also die häufigen Interaktionen erklärbar. Insofern kommt erst im zweiten Lebenshalbjahr dem Rang hohe Bedeutung zu, waren doch im ersten Lebenshalbjahr auch Kinder rangniedriger Mütter für das Alpha- Weibchen attraktiv.
62Dem Rang kommt auch bei den Interaktionen der Jungtiere zu adulten Weibchen Bedeutung zu, so interagieren die Jungen nicht nur bevorzugt mit den Sozialpartnern der Mutter - also automatisch mit Tieren des eigenen Rangbereiches, da adulte Weibchen ihre Sozialpartner im gleichen Rangbereich wählen - , sie interagieren darüber hinaus nur dann mit weiteren Weibchen, die für ihre Mütter unattraktiv sind, wenn diese rangniedriger sind als die eigene Mutter. Hier können sie sich offensichtlich annähern, ohne Abwehr befürchten zu müssen.
63Eine Ausnahme bildet hier nur ein weibliches Jungtier, das sowohl mit dem Alpha-Männchen als auch mit dem Alpha-Weibchen mehr interagierte als zu erwarten. Hierfür verantwortlich dürfte der Rang der Mutter sein, die als ranghöchste Mutter der acht untersuchten Jungtiere auch selbst häufig mit den ranghöchsten Tieren der Gruppe interagierte.
64Ausnahmen von dieser Regel waren selten. In solchen Fällen, wo die Jungtiere mit Weibchen interagierten, mit denen ihre Mütter keine Kontakte hatten, war das gewählte Weibchen stets rangniedriger als die Mutter des entsprechenden Jungtieres.
65Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1987 publizierten Beitrag ([286]).
66Da einige Individuen während der Versuche getrennt wurden (vgl. Unterkapitel 14.3), lag nicht für alle möglichen Zweierbeziehungen die gleiche Anzahl von Beobachtungsstunden vor, daher ermittelten wir die Interaktionen pro Stunde für jede einzelne Zweierbeziehung getrennt nach positiv-aktiven, positiv-passiven, positiv-neutralen und agonistischen Interaktionen. Dabei werteten wir die Daten getrennt nach Lebensjahren aus und errechneten für jedes Lebensjahr die entsprechenden Mittelwerte pro Stunde. Bedingt durch die unterschiedlichen Zeitpunkte der Geburten und die 25 Monate umfassende Beobachtungsdauer liegen Daten für die einzelnen Lebensjahre nur teilweise vor. Hier haben wir nur solche „Lebensjahre“ berücksichtigt, von denen zumindest vier Lebensmonate protokolliert wurden. Entsprechend liegen je nach Geburtsdatum die Daten für ein (bei während der Versuche geborenen Jungtieren), zwei oder drei Lebensjahren vor. Durch die Auswertung nach Lebensjahren konnten wir von den 23 für diese Untersuchung zur Verfügung stehenden Jungtieren insgesamt 965 dyadische Beziehungen analysieren. Im Befundteil (Unterkapitel 14.4.5) hatten wir durchgängig zwischen (über die Mutter) verwandten und nichtverwandten Individuen differenziert. Von den 965 Dyaden waren 39 Geschwisterdyaden. Insofern gingen in die generelle Auswertung 926 Zweierbeziehungen ein.
67Es sei hier betont, dass bei der Darstellung der Ergebnisse die absolute Häufigkeit der Kontakte nicht berücksichtigt wird, die Angaben zur Präferenz für Individuen eines bestimmten Geschlechtes sind jeweils relativ zu sehen zu den gesamten Interaktionen zur jeweiligen Altersgruppe.
68In beiden Fällen handelte es sich um die Altersgruppe der 1976 geborenen Jungtiere, die problematisch zusammengesetzt war. Von den insgesamt vier 1976 geborenen Jungtieren wuchsen nämlich nur zwei Männchen (Berni, Orbi) natürlich in der Gruppe auf, das einzige Weibchen (Birgit) und das dritte Männchen (Benni) hingegen mussten handaufgezogen werden und sind demnach den übrigen Individuen (alle in der Gruppe aufgezogen) nicht vergleichbar.
69Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1987 publizierten Beitrag ([280]).
70Zum Problem des rollenspezifischen Verhaltens sei daran erinnert, dass Geschlechterrollen für alle Vertreter des Genus Macaca beschrieben werden. Männchen und Weibchen verhalten sich unterschiedlich und können nach ihrem Verhalten eindeutig voneinander diskriminiert werden.
Bei dem Vergleich der Geschlechter untereinander sind signifikante Unterschiede bei der Häufigkeit der sozialen Körperpflege aufzeigbar, Weibchen putzen weitaus häufiger als Männchen, dabei vor allem adulte Weibchen und die eigenen Kinder. Insofern sind auch Weibchen bei sozialen Interaktionen mit bestimmten Männchen stets der aktivere Partner. Dieser Geschlechtsunterschied wird auch schon bei paarweiser Testung auffällig, hier zeigen Weibchen mehr soziale Körperpflege, Männchen hingegen kämpfen häufiger. Zudem spielen Männchen mehr. Generell halten Männchen weniger sozialen Kontakt als Weibchen. Männchen und Weibchen bevorzugen Juvenile des eigenen Geschlechtes.
Rollenspezifisches Verhalten ist also zweifellos beobachtbar. Offen ist nur, inwieweit dieses Verhalten dem jungen Makaken schon bei Geburt mitgegeben ist. Zur Klärung soll dieses Versuchsvorhaben ein Beitrag sein.
71Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1988 publizierten Beitrag ([287]).
72Auf der vorangehenden Seite habe ich die Häufigkeit des Umarmens (Embracing), Manipulierens (Manipulating) und der sozialen Körperpflege (Grooming) für unsere „peer“-aufgezogenen Jungtiere während der ersten drei Lebenshalbjahre ([196]) vorgestellt.
73Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1990 publizierten Beitrag ([283].)
74Es sei hier erneut betont, dass diese nicht in einer normalen Gruppe mit Individuen aller Altersbereiche lebten, vielmehr bestand die Gruppe nur aus Tieren desselben Altersbereiches.
75Auf der vorangehenden Seite habe ich die Häufigkeit der sozialen Körperpflege (Grooming) und des Kontaktsitzens der „peer“-aufgezogenen Jungtiere während des dritten bis sechsten Lebensjahres ([196]) angegeben (schraffiert = Männchen, weiß = Weibchen).
76Die ersten in der Gruppe geborenen Jungtiere wurden - vor allem von Laura - „geraubt“ und „liebevoll“ zu Tode gepflegt.
77Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1985 publizierten Beitrag ([246].)
78Der Text dieses Unterkapitels ist teilweise wortgleich mit einem 1985 publizierten Beitrag ([284]).
79Zusätzlich mussten wir Orbi separieren. Wir haben den Vorgang nicht beobachtet, wir vermuteten, dass Orbi Nikita niederkämpfte, anschließend aber von Majo entmachtet wurde. Orbi und Nikita haben wir dann separiert im gleichen Raum gehalten.
80Nikita zeigte fortan geradezu panisches Angstverhalten vor großen Männchen, ihm war das jahrelang gezeigte Selbstvertrauen verloren gegangen.
81Unabhängig davon, dass Majo eindeutig der Sieger war, hielt ich es wegen der unzähligen Verletzungen für geboten, ihn für einige Wochen zu separieren. Nach dem Ausheilen der Wunden durfte er in seine Sozialgruppe zurück und übernahm sofort ohne jegliche Auseinandersetzungen die Alpha-Position.
82Nikita jedenfalls blieb bis zu seinem Alterstod am 12.06.1987 ein für uns wichtiger nichtmenschlicher Mitarbeiter.
83Da ich in der Primatenstation gemeinsam mit meinen menschlichen Mitarbeitern regelmässig - letzlich ohne bleibenden Erfolg - Schabenvernichtungsaktionen durchführte, kann ich berichten, dass wir durch Schabengift keinen einzigen Primaten verloren haben.
84Diesem Vorschlag bin ich aber nicht gefolgt.
851991 hatte ich während der Tagung der International Ethological Society, auf der ich über unsere Hormonergebnisse (vgl. Unterkapitel 14.11) referierte, eine russische Primatologin, Marina Butovskaya, getroffen und mit ihr unsere Makakenergebnisse diskutiert. In Kassel lebten unter identischen „ökologischen“ Bedingungen Sozialgruppen des Javanermakaken, die sich in ihrem Verhalten auffällig unterschieden. Wie zwei verschiedene Arten demonstrierten sie unterschiedliche Sozialsysteme. Ich lud sie ein, sich davon in Kassel selbst zu überzeugen und warb auch für Marina Butovskaya ein DFG-Stipendium ein. So erhob sie im Winter 1992/1993 (also vor der Befunderhebung, von der ich hier berichtet habe) Daten an der Majo- und der Theo-Gruppe ([14]). Während dieser Zeit war die Rangordnung in der Majo-Gruppe identisch mit der, die ich oben beschrieben habe, in der Theo-Gruppe hatten nach ihrer Analyse Julia und Elke höhere Rangplätze, direkt nach Sonja und Maria, wobei Marina Butovskaya ihre Analysen auf das zu beobachtende passive agonistische Verhalten stützte, auf das ich hier nicht eingegangen bin. Grundsätzlich und erwartbar bestätigte Marina Butovskaya unsere eigenen Befunde: „Two groups of captive macaques (M. fascicularis) were studied at Kassel University, Germany. One included animals whose mothers were high-ranking, another, those whose mothers were low-ranking. The first group was a despotic community in which conflicts were severe and occurred mainly between single individuals; the reconciliation tendency was weak, the male leader was the controlling animal, and the affiliative preferences were marked. The second group was an egalitarian community split into two mutually hostile coalitions; the conflicts were less severe, the tendency for reconciliation was strong, the male leader could control only his own bloc and had no strong affiliative ties with other group members“ ([14], Seite 261).
86Ich hatte den Eindruck, sie versuchten engste Beziehungen zu dem Alpha-Männchen zu etablieren, um danach „zuzuschlagen“. Nur ging diese Rechnung nicht auf, unterstützte Majo doch stets Bedrängte und nicht Angreifer.
87„Majo played the role of the controlling animal. He was independent and self-confident, and he always helped victims irrespective of his personal preferences. Theo, on contrast, was unable to perform the controlling function, ...“ ([14], Seite 268).
88„Females born of high-ranking mothers ... were self-confident and evidently predisposed to social independence and initiative. ... Females born by low-ranking mothers ... were more cautious and, although being more often engaged in minor conflicts, never resorted to extreme forms of aggression despite the weakness and passivity of their male leader.“([14], Seite 268)
89„... our results suggest that predisposition to different social strategies is at least partly due to genetical factors.“ ([14], Seite 269).
90Majo starb wahrscheinlich den Alterstod, er war aber nur 4470 g schwer, also sichtlich stark abgemagert, so dass ich - gerade im Hinblick auf das Krankheitsbild von Theo (s. u.) nicht ausschließen möchte, dass bei ihm möglicherweise eine Diabetes-Erkrankung für den Tod mitverantwortlich war.
91Der genaue Todestag ist mit der Primatenstation verloren gegangen.
92Mir wurde bei diesen Beobachtungen schlagartig klar, warum Männchen bei vielen gesellig lebenden Primatenarten soviel größer sein müssen als ihre Weibchen. Nur große Männchen können den Weibchen standhalten, die Selektion begünstigt also größere Männchen.
93„The data of Wu et al. (1980) have been taken as convincing evidence for a gen-based mechanism that at least partially mediates kinship behavior in primates ( ...). The results of our study are at complete odds with this earlier work. We believe that this discrepancy is due to Type I statistical error in the earlier findings.“ ([58], Seite 34).
94Results indicate a very high intraclass correlation between 1/2 sibs (...) consistent with a model of complete heritability. ... The results indicate that social dominance in small groups of pre-pubescent rhesus monkeys is strongly influenced by genetic factors, and these factors are not simply artifacts of weight, sex or relative age.“ ([193]).
95„The data strongly suggests that genetic variables powerfully influence social dominance in small groups of socially housed rhesus monkeys. ... This paper suggests a large amount of the factors which influence social dominance in rhesus monkeys may be inherited. The fact that demographic variables such as age, sex and weight are not sufficient to account for these genetic influences, and the previously noted relationships between dominance and stress responses suggests dominance may be substantially determined by psychological characteristics which are themselves highly heritable. ... Nonetheless, ..., the conclusion of this experiment is that heritability strongly influences dominance rank in small groups of rhesus monkeys, and such effects are not simply artifacts of sex, age or size.“ ([193]).
96ACTH ist die Abkürzung für Adrenocorticotropes Hormon, ein in dem vorderen Anteil der Hirnanhangsdrüse, dem Hypophysenvorderlappen, produziertem Hormon, das auf die Nebenierenrinde einwirkt (Hypophyse-Nebennierenrinde-Achse). Hier bewirkt ACTH die Freisetzung des Cortisols, das wiederum in Form einer negativen Rückkoppelung die weitere Freisetzung von ACTH hemmt.
97„The results showed that higher ranking animals differed both behaviorally and physiologically from their lower ranking peers. ... Social dominance was negatively correlated with ACTH, but not significantly correlated with plasma cortisol. Consistent with this, ACTH and plasma cortisol showed no significant intercorrelation ... When the animals used in this experiment were grouped into pair of paternal 1/2 sibs and ACTH values were compared by ANOVA, significant differences were found. Additionally, these differences corresponded to the association previously found between paternity and social dominance ([193]). Infants sirred by fathers with a history of dominant offspring were showed significantly lower ACTH levels than infant sirred by fathers with a history of subordinate offspring, suggesting that both the tendency towards social dominance and high or low physiological response to stressors has significant genetic components“ ([192]).
98Adrenalin und Noradrenalin werden von Drüsenzellen des Nebenierenmarks gebildet.
99Glücklicherweise hatte ich vor dieser Entscheidung nicht eine Publikation gelesen, nach der Javanermakaken - im Gegensatz zu Rhesusaffen - als untrainierbar gelten.
100Es muss erwähnt werden, dass alle Tiere gewöhnt waren, nach einmaligem Klatschen der Hände blitzschnell das Innengehege bzw. das Außengehege aufzusuchen, um die Schieber schließen zu können. Hier kooperierten alle Makaken bei allen meiner Mitarbeiter. Das Ein- und Aussperren der Kapuzineraffen hingegen war komplizierter, hier benötigten meine Mitarbeiter einen Wasserschlauch, da einige Kapuzineraffen gerne blitzschnell im letzten Moment noch zurück in das zu leerende Gehege drängten. Das Bedrängt- und Gescheuchtwerden machte ihnen offensichtlich Freude. Nur mir gelang es, mit einer leisen Aufforderung „Geht bitte rein“, die Kapuziner ohne Stress für den Menschen einzusperren. Ich muss aber gestehen, dass ich dann, wenn irgendein Kapuzineraffe nicht das Innengehege aufsuchte, die Schieber schloß und ebenfalls leise sagte „Dann eben nicht.“. Anschließend holte auch ich den Wasserschlauch und spritzte die nicht kooperierenden Kapuzineraffen richtig nass, brachte den Schlauch wieder zurück und wiederholte meine leise Bitte. Dies merkten sich die Kapuzineraffen. Es machten ihnen Freude - trocken - zwischen meinen Mitarbeitern, die sie einsperren mussten, herumzuspringen, richtig nass werden, wollten sie aber nicht.
101Bei Mecki und Alba setzten wir eine Braunüle ein, beide Weibchen lagen eine Stunde lang widerstandslos und ruhig im Gang und - so unser Eindruck - genossen das kontinuierliche Handling.
102Bei den typischen Lernversuchen mit Futterbelohnung einzeln gehaltener Tiere geht dem Lernversuch eine mehr oder weniger „definierte“ Hungerphase zur Motivationssteigerung voraus, diese „Versuchsvorbereitung“ mag man für ethisch vertretbar halten, doch scheidet ein solches Vorgehen bei gesellig gehaltenen Individuen aus.
103Der einzige Unterschied zwischen Blutentnahmen an betäubten und wachen Individuen war die zur Blutentnahme genutzte Vene, bei betäubten Tieren wird das Blut aus einer Oberschenkelvene, bei unbetäubten aus einer Unterschenkelvene entnommen, was zweifellos schwieriger ist. Doch haben unsere Makaken die Prozedur nicht als belastend empfunden.
104Eine zusätzliche Komplikation soll noch erwähnt werden: Die sich im Gehege B aufhaltenden Javanermakaken lernten während des ersten Trainingstages, dass man von B aus den Tunnel bewegen und so das Experiment beenden kann, ohne den Tunnel aufzusuchen. Daher mussten wir die restlichen Gruppenmitglieder in A absperren. Für die letztendliche Prozedur bedeutete es, alle Individuen mussten von A nach B wechseln, ein Individuum hatte den Tunnel zu betreten und die restlichen Gruppenmitglieder mussten nach A zurückkehren. Dieses Vorgehen musste wiederholt werden, bis das letzte Gruppenmitglied den Tunnel betreten hatte.
105In Kapitel 8 habe ich ausführlich über unsere Probleme bei der Bekämpfung einer Nematoden-Infektion berichtet, hier hatte Norbert Fiege geraten, den Parasiten durch Fenbendazol zu kontrollieren.
106Nach unser Einschätzung verstand z. B. Majo, das Alpha-Männchen der ranghohen Individuuen, die Aufgabe sofort, seine Persönlichkeit gestattete es aber nicht, einfach mitzumachen, wie ein ungeplantes Experiment zeigte: Um sicher zu sein, dass jedes Individuum die gleiche Menge Brei erhält, portionierten wir diesen mit einem Plastikdosierlöffel. Majo gelang es, den Dosierlöffel zu greifen, und schleppte den gefüllten Dosierlöffel mit sich bis zum Ende der Prozedur. Nach Abschluss unseres Trainings, alle Affen waren wieder in ihren Ausgangsgehegen, setzten Annette Klaiber-Schuh und ich mich vor dem Majo-Gehege auf die Beobachterbank. Majo suchte ein höher gelegenes Ruhebrett auf, sass direkt vor uns am Vordergitter und beobachtete uns kontinuierlich. Dann - unter ständiger optischer Kontrolle der Beobachter - führte er den gefüllten Dosierlöffel zum Mund und begann Stück für Stück das Plastik abzubeißen (und dann auszuspucken), denn Brei ließ er völlig unangerührt, was sicherlich kein leichtes Unterfangen war. Frau Klaiber-Schuh und ich waren überzeugt, Majo wollte uns seine Verachtung demonstrieren.
107Sophie und Ilse waren Schwestern.
108Die Ergebnisse sind in der diesem Unterkapitel zugrunde liegenden Publikation durch Abbildungen belegt ([114]).
109Ein gutes Beispiel für die unterschiedliche Motivation der Individuen belegte Laura. Laura musste vor Versuchsbeginn aus der Majo-Gruppe entfernt werden und konnte am Training erst ab dem 15. Tag der Lernphase 2 teilnehmen. Laura wurde zuvor einzeln gehalten, erhielt aber auch die Breiportion - wie die anderen am Versuch beteiligten Tiere. Laura nahm den Brei aus unserer Hand und aß ihn sofort auf. Am ersten Tag im Tunnel aß Laura den Brei sofort. Am nächsten Versuchstag aber verschmähte sie diesen, sie wollte - wie die anderen Makaken - nur aus dem Tunnel heraus. Unabhängig davon, dass sie über Wochen gelernt hatte, dass der Brei ein begehrenswertes Gut ist, benötigte sie zehn weitere Trainingstage um das „Essen ohne Verzögerung“ wieder zu praktizieren.
110Wir haben hier nicht die Ergebnisse der beteiligten Jungtiere angegeben, die eine solche Annahme (Alter) eindeutig falsifizierten.
111Zum Einsatz unseres Trainingprogrammes ist es wichtig zu wissen, dass die zeitweise Unterbrechung des Trainings keinen negativen Einfluss auf den Trainingserfolg hat. Einmal gelernt, erinnern sich die Makaken sofort an das Vorgehen. Selbst negative Erfahrungen - das Anbieten von Metronidazol im Brei - hatte keinen negativen Einfluss auf das spätere Kooperationsverhalten.